Introduction
Stylogaster Macquart (Diptera: Conopidae) is a diverse genus of parasitoid flies in the family Conopidae (Camras and Parrillo Reference Camras and Parrillo1985). They are most prevalent in the Neotropical and Afrotropical regions and until recently only two species were known to inhabit the Nearctic Region, S. biannulata (Say) and S. neglecta Williston (Camras and Parrillo Reference Camras and Parrillo1985). The range of these two species extends from southern Ontario, Canada to Mexico (Camras and Parrillo Reference Camras and Parrillo1985). Though sharing much of the same habitat and appearing morphologically similar, they are not particularly closely related according to molecular and morphological analyses (Gibson and Skevington Reference Gibson and Skevington2013; Gibson et al. Reference Gibson, Skevington and Kelso2013).
These two species are most easily distinguished from each other by examination of the antenna. In S. neglecta the flagellum is nearly twice as long as the pedicel, while in S. biannulata the flagellum and pedicel are about equal length. Additionally, the abdomens of females of both species are distinctive; S. biannulata is more stout and short, while S. neglecta is longer and more slender (Camras and Parrillo Reference Camras and Parrillo1985).
Stylogaster has a parasitoid life history with females injecting eggs into a variety of hosts, including crickets, cockroaches (Kotrba Reference Kotrba1997; Woodley and Judd Reference Woodley and Judd1998), and possibly calyptrate Diptera (Smith Reference Smith1967; Couri and Pont Reference Couri and Pont2006; Couri and Silva Barros Reference Couri and Silva Barros2010). The latter are suspected to be accidental ovipositions, as larvae have not been reared from flies (Stuke Reference Stuke2012). Many Stylogaster species in Africa and the Neotropics are facultative or obligate army ant followers (Bequaert Reference Bequaert1922; Aldrich Reference Aldrich1930; Lopes Reference Lopes1937; Rettenmeyer Reference Rettenmeyer1961). The flies occur in diversity and numbers around the leading edge of ant swarms where they attack fleeing Orthoptera (personal observation).
Kotrba (Reference Kotrba1997) explored the possible methods of egg delivery and concluded that it is most likely that the eggs are stabbed into the host abdomen and are held there by elaborate barbs found on the eggs. Lopes (Reference Lopes1937) first described the eggs of Stylogaster and noted that many of the eggs revealed species-specific features. As a result, the eggs of all three Nearctic species are described and illustrated below.
When not attending army ant swarms, Stylogaster adults may typically be found hovering over paths in forest understorey or nectaring on small white or yellow flowers such as fennel (Foeniculum vulgare Miller (Apiaceae)), mints (Lamiaceae), and daisies (Asteraceae) (Smith and Peterson Reference Smith and Peterson1987). In parts of their range where there are no army ants, these are typically the only places to find these flies. They are relatively common but can be hard to spot (personal observation).
Materials and methods
Specimens were obtained from the following collections: American Museum of Natural History, New York, New York, United States of America (AMNH); Canadian National Collection of Insects, Arachnids, and Nematodes, Ottawa, Ontario, Canada (CNC); the University of Guelph Insect Collection, Guelph, Ontario, Canada (DEBU); the Los Angeles County Museum of Natural History, Los Angeles, California, United States of America (LACM); and the Museum of Comparative Zoology, Cambridge, Massachusetts, United States of America (MCZ). Morphological terminology follows Cumming and Wood (Reference Cumming and Wood2009) and Gibson et al. (Reference Gibson, Skevington and Kelso2013). Preliminary identifications relied on the key provided by Camras and Parrillo (Reference Camras and Parrillo1985).
Exemplar specimens were photographed with a Canon 50D EOS Digital Camera with an MP-E 65 mm macro lens (Canon Canada Inc., Mississauga, Ontario, Canada) and images were then stacked using Zerene Stacker Version 1.04 (Zerene Systems, Richland, Washington, United States of America). These images were further manipulated using Photoshop CS4 software (Adobe Systems Incorporated, San Jose, California, United States of America). Figure plates were prepared in Adobe Illustrator CS4 (Adobe Systems Incorporated). Illustrations of specimens were made by T.O.B., and were based on both lateral photographs and specimens under a dissecting scope. The microscope used was a Nikon SMZ1500 with a HI-150 High Intensity Illuminator (Nikon Canada Inc., Mississauga, Ontario, Canada).
Genitalia were excised by removing the whole abdomen, placing them in standard 1.5 mL Eppendorf tubes (Eppendorf Canada, Mississauga, Ontario, Canada) and immersing them in 95% lactic acid for no less than four hours on a 95 °C dry bath incubator (Fisher Scientific, Toronto, Ontario, Canada). Macerated abdomens were then placed on a glass depression slide and immersed in glycerin for dissection. The genitalia were then separated from the abdomen for comparison under the dissecting scope and were ultimately stored in genitalia vials with the specimens. Drawings were made of the genitalia of exemplar males, females, and eggs. All specimens examined were labelled with unique voucher numbers using the format CNC Diptera 12345 or Jeff Skevington Specimen #12345 (shortened to CNCDn and JSSn throughout the text). These numbers are cited in the material examined sections and figure captions for illustrations to allow for repeatability of the research.
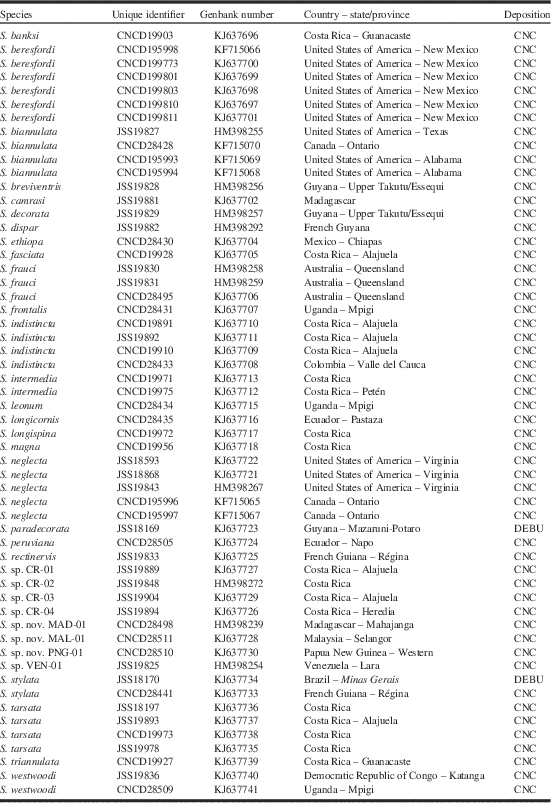
Molecular methods and analysis follow Skevington and Thompson (Reference Skevington and Thompson2012). Genetic work was carried out at the molecular laboratories of the CNC in Ottawa, Ontario, Canada and at University of Guelph, Guelph, Ontario, Canada. The resultant sequences, as well as images and related data, can be accessed through the Barcode of Life Data Systems (BOLD) (http://www.barcodinglife.org/) in the public data set “Nearctic Stylogaster (NESTY)” (http://www.boldsystems.org/views/projectmenu.php?&). In addition, all sequences were deposited in GenBank (Table 1).
Table 1 List of taxa with specimen data used in neighbour-joining tree and Bayesian analysis.

No insertions or deletions exist in the COI data set so alignment was unambiguous. Phenetic analyses were performed with PAUP* (Swofford Reference Swofford2001). Neighbour-joining was used to explore species concepts for all 35 ingroup taxa. Bayesian methods were used with a reduced data set (one exemplar per species) for creating phylogenetic hypotheses. Exemplars from all four other conopid subfamilies were used as outgroups for analyses (Dalmannia pacifica Banks, Myopa clausa Loew, Physocephala burgessi (Williston), and Zodion obliquefasciatum (Macquart)). For Bayesian Analysis, models of evolution were determined based on the Akaike Information Criterion using ModelTest 3.7 (Posada and Crandall Reference Posada and Crandall1998). Bayesian analyses were conducted using MrBayes 3.1.2 (Ronquist and Huelsenbeck Reference Ronquist and Huelsenbeck2003) with a Markov Chain Monte Carlo method. Four chains (three hot, one cold) were run simultaneously for 10 000 000 generations. Trees were sampled every 1000 generations and each simulation was run twice. Following the discard of the first 1000 samples as burn-in, 9000 samples were used for each simulation to generate a majority-rule consensus tree and posterior probabilities for each node. Analysis was conducted using the Cyberinfrastructure for Phylogenetic Research (Miller et al. Reference Miller, Pfeiffer and Schwartz2010) and a cladogram was generated using Mesquite (Maddison and Maddison Reference Maddison and Maddison2011).
Results and discussion
While testing species concepts for Nearctic conopids with mitochondrial DNA data (cytochrome oxidase c subunit 1), we discovered a putative cryptic species within S. neglecta. Upon closer examination, we found subtle, stable morphological differences between this putative new species and S. neglecta. For example, the ocellar triangle and frontal vitta is always darker, almost black, in the new species, where it is rather pale, yellow to brown, in S. neglecta. Camras and Parrillo (Reference Camras and Parrillo1985) described the triangle as dull in S. neglecta. The scape of S. beresfordi new species is always darker than S. neglecta and shiny. Upon dissection of the male and female genitalia we found other differences between these species lending support to our hypothesis.
Separating S. neglecta and S. beresfordi from S. biannulata is trivial and can be done quickly by examination of the antennae. Male and female genitalia and eggs also differ considerably between these species. We provide species descriptions, illustrations and a dichotomous key to species of Nearctic Stylogaster below.
Fig. 1 Stylogaster beresfordi new species. (A) Right lateral habitus drawing, male. Scale bar=1 mm; (B) right lateral habitus photograph, male; (C) right lateral habitus photograph, female. Photos: David Moll.
Fig. 2 Stylogaster beresfordi new species. (A) Dorsal head with antennae. Scale bar=1.8 mm; (B) left lateral habitus of abdominal ovipositor of female. Scale bar=1 mm; (C) right lateral habitus photograph CNCD 199772. Scale bar=1 mm; (D) right lateral of egg. Scale bar=0.25 mm; (E) dorsal habitus photograph CNCD 199772. Scale bar=1 mm; (F) left lateral habitus of female terminalia: cerc, cerci; lat l-st, lateral lobes of sternite 8; hyp, hypoproct. Scale bar=1 mm.
Fig. 3 Stylogaster biannulata. (A) Dorsal head with antennae. Scale bar=1.8 mm; (B) dorsal habitus of thorax. Scale bar=1.8 mm; (C) right lateral of egg. Scale bar=0.25 mm; (D) left lateral habitus of female terminalia: cerc, cerci; lat l-st, lateral lobes of sternite 8; hyp, hypoproct. Scale bar=1 mm; (E) left lateral habitus of abdominal stylus of female. Scale bar=1mm; (F) right lateral habitus photograph CNCD 195559. Scale bar=1 mm; (G) dorsal habitus photograph CNCD 195559. Scale bar=1 mm.
Fig. 4 Stylogaster neglecta. (A) Dorsal head with antennae. Scale bar=1.8 mm; (B) dorsal habitus of thorax. Scale bar=1.8 mm; (C) right lateral of egg. Scale bar=0.25 mm; (D) left lateral habitus of female terminalia: cerc, cerci; lat l-st, lateral lobes of sternite 8; hyp, hypoproct. Scale bar=1 mm; (E) right lateral habitus photograph CNCD 79778. Scale bar=1 mm; (F) dorsal habitus photograph CNCD 79778. Scale bar=1 mm.
Fig. 5 Left lateral of male terminalia. (A) Stylogaster beresfordi; (B) Stylogaster neglecta: cerc, cerci; ej apod, ejaculatory apodeme; epand, epandrium; hyp, hypandrium; ph, phallus; phapod, phallapodeme; pgt, postgonite; spm pmp, sperm pump; sur surstylus. Scale bars=0.25 mm.
Fig. 6 (A) Left lateral of male genitalia of Stylogaster biannulata: cerc, cerci; ej apod, ejaculatory apodeme; epand, epandrium; hyp, hypandrium; ph, phallus; phapod, phallapodeme; pgt, postgonite; spm pmp, sperm pump; sur, surstylus. Scale bars=0.25 mm. (B) Range map; Stylogaster beresfordi (▲), Stylogaster biannulata (●), Stylogaster neglecta (■).
Stylogaster beresfordi Burt, Skevington, and Rocha, new species
Material Examined: Holotype ♂, United States of America: New Mexico: Grant County, 14 miles North of Silver City, Cherry Creek Campground, 32.91334°N, 108.22667°W, 2256m, 16–19.viii.1999, J.E. O’Hara, Malaise trap, CNCD195998 (CNC). Paratypes: United States of America: Arizona: Santa Cruz county, Patagonia, 31.53°N, 110.57°W, 5.viii.1995, 1♀, 13.viii.1995, 1♀, 20.viii.1995, 1♂, 27.viii.1995, 1♀, B. Brown and E. Wilk. MT, LACM ENT 319810, 319811, 319812, 319813 (LACM); Oak Creek County, viii.1947, 1♂, RMB Bohart, LACM ENT 319805 (LACM). New Mexico: same location as holotype, 15–16.viii.1993, 1♂, 16–19.viii.1999, 3♂, 18–19.viii.1999, 1♀, CNCD199772–4, 199801, 199803 (CNC); Manzano Mountains, 8 miles west Manzano, New Canyon Campground, 34.67167°N, 106.41°W, 2377m, 21–23.viii.1999, 3♂, 2♀, J.E. O’Hara, Malaise trap, CNCD199802, 199810, 199811, 199812 (CNC). Mexico: Chiapas: Yerba Bueno, Highway 195, 16.93651°N, 92.89683°W, 24.vi.1969, 1♂, B.V. Peterson, abdomen missing, CNCD199744 (CNC). Sinaloa: 15 miles West of El Palmito, 23.56142°N, 105.83710°W, 1524m, 20.vii.1964, 1♂, W.R.M. Mason, CNCD199742 (CNC). Other material examined: United States of America: Arizona: Yavapai County, Bradshaw Ranger District, Prescott National Forest, Along Lynx Creek above Lynx Lake, 34.5298°N, 112.3842°W, 1695m, 12.vii.2013, 1♂, 1♀, D.D. Moll, photograph of male, sight record of female (Fig. 1C; BugGuide: http://bugguide.net/node/view/826953/bgimage).
Etymology:S. beresfordi is named after Dr. David Beresford, an entomology professor at Trent University in Peterborough, Ontario, Canada, who is a dipterist specialising in biting flies and an important mentor of the senior author.
Diagnosis: Distinguished from S. biannulata by the subequal length of the flagellum and pedicel; the flagellum is nearly twice as long as the pedicel (Fig. 2A); the ocellar triangle and frontal vitta is always dark in colour (black or dark brown) (Figs. 2A, E), darker than in S. neglecta; lateral and ventral setae on first abdominal tergite are usually pale, though this is somewhat variable (generally black in S. neglecta); the male surstylus and postgonite are both broader and stubbier than in S. neglecta (Fig. 5A).
Description:Male: Body length 6–7 mm.
Head: Face pale yellow with faint silver microtomentum extending narrowly between eyes and frontal vitta, with six to seven black setae in a row along inside of each eye; facial carina sharp and narrow; ocellar triangle with rounded protruding ocellar protuberance bearing two long setae; two long setae just posterior to protuberance; frontal vitta and ocellar triangle shiny and dark, brownish black surrounded by dull and darker V-shaped area runs along inside of eyes above antenna; dorsal occiput with distinctive patches of silver microtomentum, and long white setulae; scape, pedicel and flagellum mainly yellow, with only slight brown darkening ventrally; arista wholly dark brown; flagellum twice length of pedicel; scape appears slightly darker than in S. neglecta; subtle light-coloured microtomentum on pedicel and flagellum.
Thorax: Laterally pale yellow with brown maculation on anepisternum and katepisternum; one long black seta on anepimeron next to one shorter seta variably present; single long pale seta on lateral prosternum; thorax dorsally longer than wide (Fig. 2E); dorsal anterior postpronutum paler with pale microtomentum and single long black seta; dorsal thorax mainly black with slightly lighter brown area posteriorly and two very faint dark brown median stripes; dorsal thorax covered with short setulae only on area of darker maculation; four long black setae (not including setae on postpronutum) fairly evenly spaced along each side of dorsal lateral edge of thorax posteriorly while two postallar callus setae closer together; scutellum dark brown to black with two long posterior black setae, lighter than rest of dorsal surface of thorax.
Legs: Coxae pale yellow with hind coxa slightly darker; front coxa with one to two short black setae among pale setae; hind coxa has one longer seta laterally with one to two more medioventrally; mid coxa with several short pale and sparse black setulae anteriorly; hind legs covered in short black setulae, fore and mid legs mainly covered in pale setulae with only a few darker setulae interspersed becoming more regularly dark after tibia; fore and mid femora pale yellow; hind femur pale yellow with three darker brown bands (Figs. 1A–C, 2C), final band appearing at junction with tibia; fore and mid tibiae and tarsi pale yellow with dark setulae, hind tibia mostly pale, becoming abruptly darker brown basally, colour extending to tarsi; hind tibia with three black basal spurs; fore and mid tibiae with single pale spur; tarsomere 2 about  ${\scale60%{\vskip-7pt3}{\hskip-3pt\vskip-5.6pt\rot160/}{\scale60%{\hskip-4pt\vskip0pt4}$
length of tarsomere 1 while tarsomere 3 is about
${\scale60%{\vskip-7pt3}{\hskip-3pt\vskip-5.6pt\rot160/}{\scale60%{\hskip-4pt\vskip0pt4}$
length of tarsomere 1 while tarsomere 3 is about 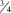 ${\scale60%{\vskip-7pt3}{\hskip-3pt\vskip-5.6pt\rot160/}{\scale60%{\hskip-4pt\vskip0pt4}$
length of tarsomere 2; tarsomere 4 and 5 equal to each other but each shorter than tarsomere 3.
${\scale60%{\vskip-7pt3}{\hskip-3pt\vskip-5.6pt\rot160/}{\scale60%{\hskip-4pt\vskip0pt4}$
length of tarsomere 2; tarsomere 4 and 5 equal to each other but each shorter than tarsomere 3.
Wing: Costal vein covered in short setulae with one long black seta dorsobasally.
Abdomen: More bulbous posteriorly (Fig. 2G); tergites pale yellow with darker brown banding and faintly darker secondary banding within, only noticeable dorsally at posterior edge of each tergite, tergites 4 and 5 darker overall with yellow banding slightly interrupted dorsally by darker maculation; tergite 2 with most prominent darker band on and towards posterior edge; ventral sternites absent except for 5th, which bears sharp carina and reduced overall; abdomen entirely covered in short black setulae dorsally, laterally black setae alternate to paler yellow where maculation paler; lateral ventral edge of tergites have long pale setae interspersed with some black setae becoming mainly black towards the posteriorly; viewed dorsally, tergite with two long black setae extending posteriorly.
Genitalia: Epandrium bulbous, rounded, with two long, black posterior bristles amidst many black setulae; surstylus with two lobes: former with small black spines on inner face and black spines on outer face with concave groove basal-laterally; latter lobe with black setae at base; cercus double-lobed, with one lobe somewhat clubbed, while other sharper distally; dorsally; cercus with many long black setae; hypandrium flat and elongate, appearing bulbous, and irregular with a short protruding lobe extending dorsally; postgonite somewhat elongate, bulbous and rounded basally, rather curled and cup-shaped with mixture of short setulae and black spines; phallus double-stranded, light brown and elongate (Fig. 5A); phallapodeme cylindrical and elongate, more bulbous distally; sperm pump only slightly smaller than epandrium, membranous; ejaculatory apodeme very rounded, teardrop-shaped.
Female: Similar to male, except banding less clear; ovipositor virtually indistinguishable from S. neglecta (Fig. 2B), twice as long as thorax and head combined; first 5th of ovipositor pale yellow graduating to darker brown with last 5th pale yellow; setae of ovipositor sparse on darker areas becoming more regular towards posterior; setulae pale yellow to white on last 5th of tergite near terminalia.
Terminalia: Tergite 7 or stylus (segment 7) cylindrical, elongate and pale with sparse, short setae (Fig. 2B); tergite 8 cylindrical and 1/3 as long as stylus with longer, coarser setae than stylus; tergite 9 short and fused dorsally just before lateral lobes of sternite 8, which are short and rounded with crop of long, bushy setulae; hypoproct arrow-shaped, short and moderately pointed with many long setae; cercus short, with very thick and long black spines, not extending to tip of hypoproct (Fig. 2F).
Egg: Anterior tip (nonmicropylar end) sharply pointed (Fig. 2D), highly sclerotised, with pair of ventral, posterior-facing (downward) spines or barbs, slightly shorter than in S. biannulata (S. biannulata egg, Fig. 3C); barbs often encased within thin translucent film; tip in shape of arrowhead or harpoon when viewed ventrally or dorsally; body elongate, cylindrical, becoming gradually more bulbous posteriorly (micropylar end), giving egg club-shaped appearance; longitudinal slit between pair of sclerotised barbs where eversible sac originates upon inflation; viewed laterally, anterior end appears rounded, or softened blade, egg body posteriorly clavate; under high magnification surface appears sculptured, scales-like, less defined or altogether absent towards anterior tip.
Hosts: Unknown, but presumably crickets and/or cockroaches as in other known Stylogaster (Smith and Cunningham-van Someren Reference Smith and Cunningham-van Someren1985; Woodley and Judd Reference Woodley and Judd1998).
Known distribution:Stylogaster beresfordi is a southwestern Nearctic species, occurring from SW New Mexico and SE Arizona south through Mexico, between 1238–2377 m in elevation (Fig. 6B).
Comments: Bayesian analayis of COI data for 35 species of Stylogaster shows S. beresfordi to be the sister species to S. neglecta (Fig. 7). Morphologically, the two species are very similar, differing only in small differences in male genitalia and colouration. DNA evidence strongly supports our hypothesis that S. beresfordi is a valid species. Six specimens were sequenced and differ by only 0.0–0.2% and cluster together in as a monophyletic lineage in a neighbour-joining tree (Fig. 8). The uncorrected pairwise genetic distance between S. beresfordi and S. neglecta is 3.6–4.2%. The COI sequences are published in GenBank (Table 1).
Fig. 7 Marjority rule consensus Bayesian-analysis cladogram based on COI sequence data. Numbers at nodes indicate posterior probability support measures. Dalmannia, Myopa, Physoconops, and Zodion were defined as the outgroup.
Fig. 8 Neighbour-joining tree based on COI sequence data.
Stylogaster biannulata (Say)
Myopa biannulata Say, Reference Say1823: 81. Type locality: United States of America, Pennsylvania. Type depository: ANSP.
Myopa?stylata Fabricius. Wiedemann Reference Wiedemann1830: 243; Röder Reference Röder1892: 287.
Stylogaster?stylatus (Fabricius). Macquart 1843: 175; Osten Sacken Reference Osten Sacken1878: 140, 259.
Stylomyia confusa Westwood, Reference Westwood1851: 269. Type locality: Unknown. Type depository: BMNH. Synonymy by Loew 1851: 133.
Stylogaster biannulata: Williston Reference Williston1883: 93 (redescription and discussion of S. stylatus misidentification by Macquart); Williston Reference Williston1885: 394 (Conopidae redescriptions); Williston Reference Williston1893: 120 (discussion of S. stylatus misidentification by Roeder); Snow Reference Snow1903: 216 (checklist of the Diptera of Kansas, United States of America); Tucker Reference Tucker1907: 99 (checklist of insects of Kansas and Colorado, United States of America); Johnson Reference Johnson1910: 772 (checklist of the insects of New Jersey, United States of America); Johnson Reference Johnson1913: 69 (insects of Florida, United States of America); Britton Reference Britton1920: 189 (checklist of insects of Connecticut, United States of America); Johnson Reference Johnson1925: 180 (checklist of Fauna of New England, United States of America); Leonard Reference Leonard1928: 803 (checklist of the insects of New York, United States of America); Aldrich Reference Aldrich1930: 7, 10 (key to New World Stylogaster species; redescription of S. biannulata); Curran Reference Curran1942: 64 (key to New World Stylogaster species); Camras Reference Camras1965: 632 (Nearctic catalogue); Camras Reference Camras1967: 9 (notes, including documentation of 60 specimens over a swarm raid of Eciton burchelli (Westwood) (Hymenoptera: Formicidae) in Mexico); Papavero Reference Papavero1971: 13 (Neotropical catalogue); Camras and Parrillo Reference Camras and Parrillo1985: 111, 115 (key to New World species; notes); Kotrba Reference Kotrba1997: 620 (egg position); Woodley and Judd Reference Woodley and Judd1998: 658 (discovery of host, Gryllus rubens Scudder; description of egg and puparium); Gibson et al. Reference Gibson, Skevington and Kelso2013: 200, 204 (illustrations of male and female genitalia; phylogeny).
Type material examined: see Remarks section.
Material examined: Canada: Ontario: Essex County, Pelee Island, Porchuk property, 41.75°N, 82.64167°W, 28.viii-10.ix.2002, 1♂, Porchuk and Marshall, Malaise trap, debu216600 (DEBU); Vineland, 43.17080°N, 79.36400°W, 3.ix.1944, 1♀, H.R. Boyce, debu115709, (DEBU); Waterloo Region, Blair, Cruickston Creek, 43.37778°N, 80.34945°W, 3027.ix.2006, 2♂, Marshall and Cheung, debu278338, 278347 (DEBU); The Dells, 43.38194°N, 80.34223°W, elevation 265 m, 20.vi.2006, 17.vii.2006, 1♂, Bergeron and Cheung, Malaise trap, debu26874, (DEBU); Puslinch, 43.43334°N, 80.08334°W, 18–20.ix.1983, 1♀, Coote and Marshall, Malaise trap, debu1157050 (DEBU); Arkell, 43.53729°N, 80.16782°W, 21.vi.1986, 1♂, 29.vi.1986, 3♂, 2♀, E. Lippert, debu1157064, 1157070–4 (DEBU); 18.viii.1987, 1♀, 25.viii.1987, 1♂, 1♀, 30.viii.1987, 1♂, 13.ix.1987, 1♂, Bob Westhaver, Malaise trap, debu1157065–9 (DEBU); Guelph, 43.54480°N, 80.24816°W, 23.viii.1987, 1♂, Bob Westhaver, debu1157090 (DEBU). Northumberland County, Barr property, ~7 km NE Centreton, 44.13°N, 77.98416°W, 27.vi.-2.vii.2011, 1♀, 12–26.vii.2011, 1♀, Brunke and Paiero, Malaise trap, debu1152347, 1154032 (DEBU); Ferris Provincial Park, Drumlin Trail, 44.29134°N, 77.79383°W, 1–15.ix.2003, 1♀, P.D. Careless, debu1157092 (DEBU). Chaffey’s Locks, Leeds County, 44.5658°N, 76.3242°W, 31.viii.1973, 1♀, P. Ward, CNC Diptera 199681 (CNC); North Burgess Township, Lanark County, 44.80696°N, 76.31263°W, 7.ix.1970, 1♀, 19–26.vii.1971, 1♂, D.M. Wood, Malaise trap, CNCD199695, 199708 (CNC); Innisville, 45.05140°N, 76.25050°W, 12.vii.1961, 1♀, W.R.M. Mason, CNCD199687 (CNC); Metcalfe, 45.23582°N, 75.47271°W, 25.ix.1982, one abdomen missing, 30.ix.1982, 1♀, 8.ix.1983, 1♂, 2♀, 22.ix.1983, 1♂, 21.viii.1984, 1♂, 22.viii.1984, 1♂, 6.ix.1984, 2♂, 27.vi.1985, 2♀, 28.vi.1985, 1♀, 5.vii.1985, 1♀, 7.vii.1986, 2♂, 5♀, 17.vii.1986, 1♀, 11.ix.1993, 1♀, 12.vii.1993, 1♀, 1♀, 27.viii.1994, 1♂, B.E. Cooper, CNCD28428, 89252, 195995, 199713, 199702–7, 199712–6, 199721–3, 199732–5, 199793, 199797, 199805 (CNC); 2 miles North of Metcalfe, 45.26540°N, 75.48873°W, 20.iv.1952, 1♀, 25.vi.1982, 1♀, 28.vi.1982, 1♂, 1♀, 11.ix.1982, 1♂, 1♀, 14.ix.1982, 1♂, 20.ix.1982, 1♂, 21.ix.1982, 1♂, 1♀, 22.ix.1982, 1♂, 6.x.1982, 1♂, 4.vii.1984, 2♀, 6.vii.1984, 1♀, B.E. Cooper, CNCD79762–3, 89251, 199696–7, 199699–701, 199711, 199725–9, 199733 (CNC); Ottawa, near Uplands Airport, 45.33334°N, 75.58334°W, 16-viii.1937, 1♂, 6♀, 15.viii.1987, 1♂, 1♀, 6–12.vii.1989, 1♀, J.M. Cumming, Malaise trap, CNCD199698, 199792, 199787–94, 199816–7 (CNC); Dunrobin, 45.42221°N, 76.02026°W, 18.vii.1972, 3♀, H.J. Teskey, Malaise trap, CNCD199685, 199689, 199719, (CNC); Quebec, 4 miles North of Eardley, 45.60581°N, 75.87640°W, 25.viii.1971, 1♀, D.M. Wood, CNCD199686 (CNC); Masham Township, Gatineau County, 45.68334°N, 76.05°W, 6–8.vii.1974, one missing abdomen, 2.viii.1974, 1♀, 7–14.ix.1974, 1♀, 2.viii.1974, 1♀, 6–8.vii.1974, 1♀, 10–20.vii.1974, 1♀, 20–23.viii.1975, 1♀, 25–28.viii.1975, 1♀, D.M. Wood, CNCD199683, 199691, 199693, 199709, 199717–8, 199720 (CNC). Mexico: Yucatan: Chichen Itza, 20.68428°N, 88.56778°W, no/day.vi.1929, 1524m, 2♀, JSS26053, 26056 (AMNH). United States of America: Alabama: Baldwin County, Bon Secur, 30.3°N, 88.74°W, 19.x.2004, 5♀, E. Benton, Malaise trap, CNCD199736, 199738–41 (CNC); Auburn University, 30.55°N, 88.87°W, 15.xii.2004, 2♀, E. Benton, Malaise trap, CNCD195993, 199819 (CNC); Silver Hill, 30.56°N, 87.77°W, 15.xii.2004, 1♂, E. Benton, Malaise trap, CNCD195994 (CNC). Florida: Okeechobee County, Archbold Biological Station, Lake Placid, Highlands County, Trail 1 SSo, 27.188°N, 81.338°W, 9.xii.1985, 1♂, 1♀, 13.xii.1985, 2♂, 25.xii.1985, 3♂, 1♀ M. Deyrup, Malaise trap, debu1157055, 1157056–62, DEBU; Okeechobee County, Archbold Biological Station, Lake Placid, Highlands County, Trail 1 SSo, 27.18966°N, 81.33813°W, 15.xii.1985, 1♂, 18.xii.1985, 1♂, 1♀, S.A. Marshall, 14–17.iv.1989, 5♂, Malaise trap, debu1157041–3, 1157045–9 (DEBU); Okeechobee County, Archbold Biological Station, Lake Placid, Highlands County, Trail 1 SSo, 27.20736°N, 81.33813°W, 4–10.viii.1987, 1♂, 1♀, CNCD199800, 199814 (CNC); Okeechobee County, Archbold Biological Station, Lake Placid, Highlands County, Trail 1 SSo, 27.297°N, 81.37°W, 9.xii.1985, 8♂, 2♀, 20.xii.1985, 3♂, 3♀, 9.xii.1985, 1♂, 1♀, M. Deyrup, Malaise trap, debu1157052–4, 1157063, 1157075–89 (DEBU); Highlands County, Hammock State Park, 27.46667°N, 81.53334°W, 17.iv.1989, 1♀, B.J. Sinclair, 13–17.iv.1989, 1♀, J.E. Swann, Malaise trap, CNCD199710, debu1157051 (CNC, DEBU); Gainesville, 29.65°N, 82.31667°W, 24.iv.1952, 1♂, G.S. Walley, 8–22.xii.1986, 1♀, W. Mason, Malaise trap, CNCD195559, 199795 (CNC). Georgia: Pierce County, 5 km North of Bristol, 31.4935°N, 82.21492°W, 18.iv.1989, 1♂, K.N. Barber, sweep, roadside, debu1157044, (DEBU). Illinois: Chicago, 41.87811°N, 87.6298°W, 6.vii.1895, 1♀, W.M. Wheeler collection, JSS26058 (AMNH). Kansas: Manhattan, 39.18361°N, 96.57167°W, 29.x.1932, 1♀, C.W. Sabrosky, JSS26055 (AMNH). Mississippi: Winston County, Tombigbee National Forest, 33.255°N, 89.091389°W, 10.viii.1952, 1♀, J.M. Cumming, Malaise trap, CNCD222738 (CNC); Agricultural College, 33.45553°N, 88.79042°W, 8.x.1920, 1♀, 6.x.1921, 1♀, CNCD199679, 199684 (CNC); Oxford, 34.36649°N, 89.51924°W, v.1940, 3♂, 1♀, 10.vi.1945, 1♀, F.M. Hull, Frank M. Hull Collection C.N.C. 1973, CNCD79761, 85163,199682, 199688, 199692 (CNC); Tishomingo State Park, 34.59941°N, 88.18456°W, 20.v.2013, G.A. Dahlem, Malaise trap, JSS26307, CNC; Rosedale, 39.32010°N, 76.51551°W, 9.ix.1922, 1♀, 14.ix.1922, CNCD199678, 199680 (abdomen and head missing) (CNC); Williamsville, 36.97249°N, 90.54833°W, x-xi.1968, 2♂, Malaise trap, 1♂, 1♀, J.T. Becker, CNCD199690, 199694, 199730–1 (CNC). New York: Rochester, 43.16103°N, 77.61092°W, 18.ix.1933, 1♀, R.L. Post, JSS26054, AMNH; Nyack, 41.09065°N, 73.91792°W, 1883, 1♀, JSS26057 (AMNH). Ohio: Springfield, 39.92423°N, 83.80882°W, 8.ix.1937, 1♀, R.H. Crandall, JSS26059 (AMNH); South Carolina: Dorchester County, Francis Beidler Forest, 10 km Northeast of Harleyville, 33.22064°N, 80.35419°W, 28.iv.1987, 1♂, Malaise trap, CNCD199813 (CNC); Texas: Hardin County, 4 miles East of Silsbee on Route 418, Roy E. Larson Sandylands Sanctuary, 30.3°N, 88.74°W, 17–19.v.1993, 1♀, J.M. Cumming, Yellow Pan Trap and Malaise Trap, CNCD199737 (CNC); Brazos County, College Station, Lick Creek Pk., 30.56223°N, 96.21472°W, 30.iii-5.iv.2000, 1♀, 5–9.iv.2000, 1♀, M. Buck, Malaise trap, debu110561, 114275, DEBU; Lamar County, Camp Maxey, 33.86427°N, 95.50661°W, 21.x-8.xii.2003, 1♀, W. Godwin, Malaise trap, JSS19827 (CNC).
Diagnosis: Antennae with near equal lengths of pedicel and flagellum (Fig. 3A); ovipositor of female stout compared to S. neglecta and S. beresfordi (S. biannulata stylus Fig. 3E).
Redescription: Male: Body length 6–8 mm.
Head: Face pale yellow with extremely faint silver microtomentum extending narrowly between eyes and frontal vitta with four to six variably pale yellow or black setae in a row along inside each eye; facial carina sharp and narrow; ocellar triangle with slight, irregularly rounded ocellar protrusion with two setae just behind protrusion; frontal vitta and triangle are somewhat dull pale yellow near scape and becomes dark brown to black just before ocellar protrusion, entirely surrounded by a dull and darker V-shaped area (Fig. 3A); back of the head just behind eyes with distinctive patches of silver microtomentum, and long white setulae on lateral occiput; scape, pedicel and flagellum mainly yellow with darker brown on the flagellum, with feather-like arista mainly yellow, and pedicel mainly darker brown; flagellum equal in length with pedicel; scape with short subtle and sparse black pile; pedicel with moderately long, spikey black pile.
Thorax: Laterally pale yellow with light brown maculation on anepisternum and katepisternum (Fig. 3F); thorax dorsally longer than wide (Figs. 3B, G); dorsal anterior postpronutum paler with pale microtomentum and without long setae, dorsal thorax mainly black with slightly lighter brown area posteriorly; dorsal thorax covered with short pale setulae; four long black setae fairly evenly spaced along each side of the dorsal lateral edge of thorax distally, with two postallar callus setae closer together; scutellum dark brown to black with two posterior long black setae; scutellum slightly lighter than rest of dorsal surface of thorax.
Legs: Coxae pale yellow with hind coxa slightly darker; fore coxa with several long pale and black setae; hind coxa with mainly black, long and short setae, mid coxa with several long pale and sparse black setulae anteriorly; hind legs mainly covered in short black setulae except ventral surface of fore femur with longer black setulae; fore and mid legs covered in pale setulae with only a few darker setulae becoming more regularly dark after tarsomere 3; hind femur pale yellow with two darker brown bands (Figs. 3F, G); fore and mid tibiae and tarsi pale yellow setulae; while hind tibia is mostly pale, becoming more stout and broad, and abruptly darker brown basally, colour extends throughout hind tarsi (Fig. 3F); hind tibia has three fairly reduced and subtle basal spurs; fore and mid tibiae with single pale spur; tarsomere 2 about 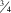 ${\scale60%{\vskip-7pt3}{\hskip-3pt\vskip-5.6pt\rot160/}{\scale60%{\hskip-4pt\vskip0pt4}$
length of tarsomere 1; tarsomere 3 is about 1/3 length of tarsomere 2; tarsomere 3–5 equal to each other; fore and mid tarsi pale yellow, except for brown 5th.
${\scale60%{\vskip-7pt3}{\hskip-3pt\vskip-5.6pt\rot160/}{\scale60%{\hskip-4pt\vskip0pt4}$
length of tarsomere 1; tarsomere 3 is about 1/3 length of tarsomere 2; tarsomere 3–5 equal to each other; fore and mid tarsi pale yellow, except for brown 5th.
Wing: Costal vein covered in short setulae with one long black seta dorsal-basally.
Abdomen: More bulbous distally; tergites mainly pale yellow with gradient darker brown banding, darker towards posterior margin of tergite, when viewed dorsally (Fig. 3G); ventral sternites mainly absent except sternite 5, which bears sharp carina, reduced overall; dorsal abdomen entirely covered in short black setulae, anteroventrally tuft of paler yellow setae present; viewed dorsally, tergite 3 appears somewhat fluted posteriorly, tergite 4 bulbous.
Terminalia: Epandrium not bulbous, rather flattened dorsally, without long, posterior bristles, with many fine black setulae; surstylus slightly L-shaped with slight second lobe at bend, distal lobe with sparse black setulae on outer face and shorter black setae on inner face (Fig. 5B); cercus single-lobed, and sickle-shaped, sharp and pointed distally; dorsally, cercus with many thin, long black setae; postgonite somewhat elongate, carnate dorsally, with short, sharp black spine-like setae ventrally only, appearing brush-like; phallus Y-shaped, single stranded later dividing into short double-stranded form, dark brown (Fig. 5B); phallapodeme cylindrical and elongate, becoming narrower distally; sperm pump only slight, much smaller than epandrium (about size of surstylus), rounded; ejaculatory apodeme is thin, somewhat narrow, leaf or arrow-shaped, thin and somewhat flat (Fig. 5B).
Female: Similar to male, except for genitalia; ovipositor shorter and stouter than in S. neglecta and S. beresfordi, with a slightly uneven lateral groove (Fig. 3E).
Genitalia: Tergite 7 or ovipositor, cylindrical, only somewhat elongate and pale with sparse, short setae, and a lateral groove, or crease (Fig. 3E); tergite 8 cylindrical and 1/3 as long as ovipositor with longer, coarser setae; tergite 9 short and fused dorsally, somewhat protruding dorsally, lateral lobes of sternite 8 with sparse setulae (Fig. 3D); hypoproct appears mildly arrow-shaped, short and stout, only moderately pointed with many medium length setae; cercus slightly elongate and sickle-shaped, with long black spines, not extending to tip of hypoproct (Fig. 3D).
Egg: The eggs of S. biannulata have been thoroughly described by Woodley and Judd (Reference Woodley and Judd1998) but are re-illustrated and described here for comparative purposes (Fig. 3C). Anterior tip (nonmicropylar end) sharply pointed and highly sclerotised with pair of ventral, posterior-facing (downward) spines or barbs, slightly longer than S. beresfordi or S. neglecta (S. beresfordi and S. neglecta eggs, Figs. 2D, 4C); barbs are often encased within thin, translucent film; tip in shaped of arrowhead or harpoon when viewed ventrally or dorsally; body elongate, cylindrical, becoming gradually more bulbous towards the posterior (micropyle end), club shaped; longitudinal slit opening between two sclerotised barbs where the eversible sac originates upon inflation; viewed laterally, anterior end appears much like a sharp and pointed blade while egg body posteriorly clavate; under high magnification surface appears sculptured, scale-like, that are less defined or altogether absent towards the posterior.
Hosts: Woodley and Judd (Reference Woodley and Judd1998) noted that Gryllus rubens Scudder serves as the host for S. biannulata. These crickets were often parasitised more than once with larvae emerging roughly 19–25 days later. Camras (Reference Camras1967) documented a collection of 60 adult specimens taken over a swarm raid of Eciton burchelli in Mexico. It thus seems likely that they also attack more than just Gryllus rubens. More work is needed to understand host use in species of Stylogaster.
Known distribution:Stylogaster biannulata occurs from southern Mexico north to Oklahoma, Kansas and Nebraska, east to Florida in the United States of America and north to Ontario and southern Québec, Canada (Fig. 6B).
Comments: Four COI sequences have been obtained and are published in GenBank (Table 1). Intraspecific variation amounted to 0.2–1.5%. The uncorrected pairwise genetic distance between S. biannulata and S. beresfordi is 15.0–21.9% and between S. biannulata and S. neglecta is 17.9–19.9%. Based on a combined five-gene and morphology phylogenetic analysis of Conopidae, S. biannulata was hypothesised to be in a different species group than S. neglecta (Gibson et al. Reference Gibson, Skevington and Kelso2013). Eleven of the 59 conopid species included in their data set were Stylogaster. We now have additional COI data available for conopids and include 35 species of Stylogaster in the Bayesian analysis presented here (Fig. 7). We recover a similar pattern of relationships to those found by Gibson et al. (Reference Gibson, Skevington and Kelso2013) despite using only COI data in the analysis.
The holotype of S. biannulata is missing and has been missing for decades. It was deposited in the Academy of Natural Sciences in Philadelphia. Until the authors have a chance to visit this collection in person it is likely not wise to nominate a neotype. Given the distinctiveness of the taxon, it also seems unnecessary to designate a neotype.
Stylogaster neglecta Williston
Stylogaster neglecta Williston, Reference Williston1883: 91. Type locality: United States of America, Connecticut. Type depository: syntypes MCZ, UKAL.
Stylogaster neglecta Williston. Williston Reference Williston1894: V4 (checklist of insects); Townsend Reference Townsend1895: 64 (checklist of Diptera of North America); Johnson Reference Johnson1925: 180 (catalogue of insects of New England, United States of America); Snow Reference Snow1903: 216 (checklist of Diptera for Kansas, United States of America); Jones Reference Jones1907: 251 (a preliminary checklist of conopids in Nebraska, United States of America); Williston Reference Williston1908: 262 (description and checklist); Banks Reference Banks1912: 109 (checklist of insects); Banks Reference Banks1916: 200 (checklist of insects of northern Virginia, United States of America); Greene Reference Greene1913: 45 (checklist and notes on natural history and behaviour); Kröber Reference Kröber1914: 349 (catalogue of insects); Britton Reference Britton1920: 189 (checklist of insects of Connecticut, United States of America); Johannsen Reference Johannsen1928: 803 (checklist of insects of New York, United States of America); Aldrich Reference Aldrich1930: 7, 10 (key to New World Stylogaster species; redescription of S. neglecta); Curran Reference Curran1942: 64 (key to New World Stylogaster species); Papavero Reference Papavero1971: 13 (Neotropical catalogue); Camras and Parrillo Reference Camras and Parrillo1985: 111, 115 (key to New World species; notes); Kotrba Reference Kotrba1997: 620 (egg position); Gibson et al. Reference Gibson, Skevington and Kelso2013: 200, 204 (illustrations of male and female genitalia; phylogeny).
Type Material Examined: Holotype ♂, [United States of America: Connecticut], Osten Sacken Coll., Type Wlstn., MCZ-ENT00000450 (MCZ). Other material examined: Canada: Ontario: Essex County, Pelee Island, Porchuk property, 41.75°N, 82.64167°W, 26.vi.-8.vii.2002, 3♂, 8.vii-14.vii.2002, 3♂, 17–22.vii.2002, 1♂, 25.vii.2002, 1♂, 1♀, 25.vii-3.viii.2002, 2♂, 2♀, B. Porchuk, Malaise trap, debu198135, 198141, 198144, 198158–60, 216482, 216484, 216486–7, 227820–2 (DEBU); Essex County, Point Pelee National Park, 41.95834°N, 82.5125°W, 19.vii.1983, 1♀, Marshall, Logan and Grigsby, 20.vii.1978, 15♂, 2♀, J.M. Cumming, 16.vii.1962, 1♂, 1♀, S.M. Clark, 17.vii.1978, 1♂, 18.vii.1978, 1♂, 31.vii.1978, 1♂, D. Morris, 18.vii1978, 2♂, W.A. Attwater, 20.vii.1978, 10♂, 22.vii.1979, 1♂, K.N. Barber, debu19035, CNCD199758–9, debu6005–18, 6023–8, 6030, 6032–6, 19032–3, 19036–7 (CNC, DEBU); Kingsville, 42.03650°N, 82.73909°W, 8.vii.1977, 4♂, 1♀, W.A. Attwater, debu1157113–5, 1157117, 1157119 (DEBU); Windsor, ~1.5 km S Ojibway Prairie, 42.22611°N, 83.07416°W, 19–30.vi.2001, 2♂, 30.vi-17.vii.2001, 1♀, P. Pratt, 18–31.vii.2001, 1♂, 2♀, S.M. Paiero, 42.29267°N, 82.99333°W, 4.vii.1984, 2♂, M.T. Kasserra, Malaise trap, debu1106076–7, 1107993–4, 1108002, 1108109, 1157094, 1157104 (DEBU); Maidstone Conservation Area, 42.21383°N, 82.78694°W, 21.vii.2000, 1♂, 1♀, S.A. Marshall, debu137396 (DEBU); Walpole, 42.61504°N, 82.51457°W, 12.vii.1979, 1♀, D. Morris, debu1157098 (DEBU); Elgin County, Fingal Wildlife Management Area, 42.67948°N, 81.32660°W, 13.viii.1997, 1♀, 29.vi.1998, 1♂, 3.viii.2000, 1♀, 27.vii.2002, 1♂, 1♀, Ian Carmichael, Newport Forest, ~3 km SW of Wardsville, 42.63111°N, 81.77861°W, 30.vii.2009, 1♂, G.F.G. Miranda, debu318212, CNCD195996–9, 199818 (CNC, DEBU); Regional Municipality of Haldimand-Norfolk, Cronmiller property, ~6 km W St. Williams, 42.6725°N, 80.49056°W, 20.vii.2011, 1♂, S.M. Paiero, debu1148553, DEBU; Manestar Tract, 6 km NNW St. Williams, 42.70472°N, 80.46056°W, 3.viii.2001, 1♀, M. Parchami-Araghi, debu166472 (DEBU); Turkey Point Provincial Park, 42.71167°N, 80.34223°W, 6–22.vii.2009, 1♀, 22.vii-3.viii.2009, 1♂, 3♀, S.M. Paiero, Malaise trap, debu336037, 1150079, 1150081, 1150085 (DEBU); Delhi Simcoe Railway, 42.85°N, 80.38334°W, 14.vii.2001, 2♂, 1♀, S.M. Paiero, sweep, debu1104992, 1106599, 1116587 (DEBU); Jordan, 43.13334°N, 79.36667°W, 13.vii.1919, H. Curran, Compared with type (of Stylogaster neglecta Williston, Reference Williston1883); missing abdomen, 4.vii.1920, 1♂, C.H. Curran, 13.vii.1919, 1, 14.vii.1919, 1♂, 1, 18.vii.1919, 1♂, 1♀, 25.vii.1919, 1♂, W.A. Ross, CNCD199770, 199778–9, 199781–3, 199785, CNC, 17.vii.1919, 1♂, W.A. Ross, debu1157108 (DEBU); Brant County, Brantford Railway Prairie, 43.16667°N, 80.31667°W, 24.vii.2002, 1♀, S.M. Paiero, debu1115762 (DEBU); Hamilton, 43.23333°N, 79.95°W, 24.vii.1982, 1♂, Kevin Barber, debu1157099 (DEBU); Pinery Park, Chalet, 43.255124°N, 81.8388297°W, 9.vii.1991, 1♂, J. Malaise trap, debu1157106 (DEBU); Dundas, 43.263453°N, 79.9527°W, 7–11.vii.1970, 1♂, 7–11.vii.1972, 4♂, 2♀, D.M. Wood, CNCD 28437 (CNC); 24.vii.1975, 1♂, D.J. McComb, 21.vii.1981, 3♂, Cyrus Farivar, debu1157100, 1157102, 1157118, 1157112 (DEBU); Waterloo Region, Blair, RARE, Cruickston Creek, 43.37778°N, 80.34945°W, 301, 27.ix.2006, 1♂, Marshall and Cheung, debu278326 (DEBU); Cambridge, 43.38334°N, 80.31667°W, 24.viii.1989, 1♀, M.K. Sears, Malaise trap, debu1157097, DEBU; Freelton, 43.39779°N, 80.03642°W, 19.viii.1984, 1♂, M.T. Kasserra, debu1157105 (DEBU); Regional Municipality of Halton, Oakville, nr. Hwy 25 and Burnhamthorpe Rd., 43.45389°N, 79.79223°W, 26.vii-16.viii.2006, 6♂, 5♀, S.M. Paiero, Malaise trap, debu285658–63, 285665–7, 285686–7 (DEBU); Bronte Creek Provincial Park, 43.415°N, 79.76667°W, 19.vii.2002, 5♂, K.N. Barber, sweep, debu1157109–11, 1157120–1 (DEBU); Guelph, 43.54480°N, 80.24816°W, 27.vi.1955, 1♂, D.H. Pengelly, debu 1157116 (DEBU); Fergus, 43.7°N, 80.36667°W, 4.vii.1990, 1♀, S.A. Marshall, Malaise trap, debu1157095 (DEBU); Bothwell, 45.05140°N, 76.25050°W, 18.vii.1962, 1, S.M. Clark, missing abdomen, CNCD199784 (CNC); 2 miles North of Metcalfe, 45.26540°N, 75.48873°W, 16.vii.1982, 1♀, B.E. Cooper, CNCD199775 (CNC). United States of America: Georgia: Rabun Bald, Rabun County, 34.964827°N, 83.298647°W, 914, 14.vii.1957, 1♂, J.G. Chillcott, CNCD199755 (CNC). Kansas: Lawrence, 38.97166°N, 95.23525°W, 22.vi.1922, 1♂, C.H. Curran, CNCD199780 (CNC). Kentucky: Manifee county, near Frenchburg, 37.98°N, 80.68°W, 2–8.vii.1997, 1♂, 15–22.vii.1997, 1♂, 2♀, Abnee and Sharkey, MT, Malaise Trap, LACM ENT 319801, 319802, 319803, 319804 (LACM). Maryland: Calvert County, Port Republic, 38.5°N, 76.53334°W, 3–5.vii.1993, 1, 8♂ and 3♀, 25.viii.2001, 1♂, D.M. Wood, 7–8.viii.1993, 1♀, Xuekui Sun, CNCD79785, 89262–3, 199762–7, 199769, 199806, debu1157101 (CNC, DEBU); Patuxent Wildlife Center, 38.53996°N, 76.74768°W, 6–22.viii.1979, 1♂, 2♀, 5.viii.1979, 1♀, E.E. Grissell and M. Schauff, CNCD199807–9, 199815 (CNC); Plummers Island, 38.97004°N, 77.17880°W, 3.vii.1921, one sex not determined, 14.vii.1921, 5♂, one sex not determined, J. Bequaert collection, JSS23095, 23096, 23097, 23098, 23099, 23100, 23101 (MCZC); 18.vii.1909, 1♀, G.P. Engelhardt, JSS23102 (MCZC). Massachusetts: Auburndale, 42.34334°N, 71.24771°W, 16.vii.1903, 1♂, CNCD199756 (CNC); Riverside, 42.36768°N, 71.11367°W, 12.vi.no/year, 1♂, 12.vi.1903, one sex not determined, A.P. Morse, JSS23103, 23104 (MCZC). Mississippi: Oxford, 34.36649°N, 89.51924°W, v.1940, 6♂, F.M. Hull, Frank M. Hull Collection C.N.C. 1973, CNCD199751–2, 199760, 199771, 199776–7 (CNC). Missouri: Williamsville, 36.97249°N, 90.54833°W, 14.vii.1969, 1♂, 22.vii.1969, 1♀, Malaise trap, CNCD199753, 199757 (CNC); La Clede County, Bennett Springs State Park, 37.72141°N, 92.85296°W, 1.viii.1984, 1♂, B.V. Brown, debu1157103 (DEBU); Rufolf Bennitt Wildlife Area, Randolf County, south of Moberly, 39.24812°N, 92.45547°W, 25.vii.1970, 1♂, R. Heitman, JSS26125, (AMNH). New Jersey, Riverton, 40.01150°N, 75.01489°W, 21.vii.1920, 1, 3♂, 1♀, Frank M. Hull Collection C.N.C. 1973, CNCD199743, 199747–50 (CNC); Alpine, 40.95593°N, 73.93124°W, 28.vii.1946, 3♂, 1♀, S.C. Harriot, JSS26085, 26086, 26087, 26088 (AMNH); Kalbfleisch Research Station, Huntington, Long Island, 40.80482°N, 73.33623°W, 9.vii.1918, 1♂, A. Nicolay, JSS26113, AMNH; Ramsey, 41.05741°N, 74.14089°W, 4.viii.1935, 1♂, JSS26114, AMNH; 40.01150°N, 75.01489°W, 15.vii.no/year, 1♀, 5.viii.no.yr, 1♂, 30.vii.1888, 2♂, 22.vii.1920, 1♂, C.W. Johnson, JSS23105, 23106, 23107, 23108, 23109, 23110 (MCZC). New York, Kalbfleisch Research Station, Huntington, Long Island, 40.80482°N, 73.33623°W, 26.vii.1962, 1♀, P.H. Arnaud, Malaise trap, CNCD199786 (CNC); Cold Spring Harbor, Long Island, 40.87148°N, 73.45678°W, 18.viii.1932, 2♂, 19.viii.1932, 2♂, 22.viii.1932, 4♂, 23.viii.1932, 8♂, 26.viii.1932, 1♂, C.H. Curran, JSS26060, 26061, 26062, 26063, 26064, 26064, 26065, 26066, 26067, 26068, 26069, 26070, 26071, 26072, 26072, 26073, 26074, 26075, 26076, 26077, 26078, 26079, 26079, 26080, 26081 (AMNH); Tuxedo, Station for the Study of Insects, 41.23766°N, 74.19445°W, 25.viii.1928, 1♂, 28.viii.1928, 1♂, 1♀, C.H. Curran, JSS26082, 26083, 26084, AMNH; 27.viii.1932, 1♂, 1♀, Ethel Curran, JSS26073, 26081, AMNH; 40.80482°N, 73.33623°W, 19.vii.1962, 1♂, 1♀, 24.vii.1962, 4♀, 26.vii.1962, 1♂, 4♀, 28.vii.1962, 4♀, 1.15.viii.1962, 3♀, P.H. Arnaud, JSS26089, 26090, 26091, 26092, 26092, 26093, 26094, 26095, 26096, 26097, 26098, 26099, 26100, 26101, 26102, 26103, 26104, 26106, 26107, 26113, 26114 (AMNH); 19.vii.1962, 1♂, Rozen, Arnaud, Hessel, Woods, Statham, JSS26105, AMNH; 19.24.vii.1976, 1♂, 19.28.vii.1976, 1♂, 25.31.vii.1976, 2♂, M. Kamran JSS26108, 26109, 26110, 26110 (AMNH); 8.18.viii.1971, 1♂, P. and B. Wygozinsky, JSS26112, AMNH; Long Island, Flatbush, water works, 40.64092°N, 73.96243°W, 11.vii.1890, 1♂, JSS26124 (AMNH); Fishers, 43.0094°N, 77.45514°W, 9.vii.1933, 3♂, 16.vii.1933, 4♂, JSS26115, 26116, 26117, 26118, 26119, 26120, 26121 (AMNH); Mosholu, 40.87700°N, 73.88489°W, (no date) 1♂, JSS26122 (AMNH); Ithaca, Tompkins County, 42.44458°N, 76.49926°W, 22.vii.1979, 1♀, 23.vii.1979, 3♂, 1♀, 3.viii.1979, 1♂, J.E. Rawling, JSS23116, 23117, 23118, 23119, 23120, 26121 (MCZC). Ohio: Lawrence County, Dean State Forest, 38.68716°N, 82.62133°W, 13.vii.1989, 1♂, S.A. Marshall, debu1157096 (DEBU). Tennessee: Sevier County, Great Smoky Mountains National Park, ATBI Plot: Twin Creeks, 35.685°N, 83.499°W, 21.vii-9.viii.1999, 1♂, Parker, Stocks, Perterson, Malaise trap, CNC Diptera 199761 (CNC). Vermont: Calisol, 18.viii.1802, 1♂, JSS26123 (AMNH). Virginia: Buffalo Mountain, 2.5 km W of Moles Road, 36.79605°N, 80.47616°W, 1211, 24.vii.2009, 1♂, J. Skevington, JSS18593 (CNC); Fish Hatchery Lane, 2.5 km S Montebello, 37.84191°N, 79.13125°W, 825m, 22.vii.2009, 4♂, J. Skevington, JSS18799, 18868–9, 19843 (CNC); Falls Church, 38.86667°N, 77.16667°W, 14.vi.no/year, 1♀, 22.vi.no/year, 1♂, 28.vi.no/year, 2♂, 17.vii.no/year, 1♂, 27.vii.no/year, 1♂, 5.viii.no/year, 1♂, N. Banks, JSS23088, 23089, 23090, 23091, 23092, 23093, 23094 (MCZC); Glencarlyn, 38.86379°N, 77.12694°W, 17.vi.no/year, 1♂, 21.vi.no/year, 1♀, 2.vii.no/year, 1♂, N. Banks collection, JSS23111, 23112, 23113 (MCZC).
Diagnosis: Distinguished from S. biannulata by the subequal flagellum and pedicel; the flagellum is nearly twice as long as the pedicel (Fig. 4A); ocellar triangle usually light in colour (tan or yellow) (Figs. 4A, F), lighter than in S. beresfordi; long ventral and lateral setae on first abdominal tergite are usually black rather than pale, though this character is somewhat variable; male surstylus and postgonite are both narrower and more elongate than in S. beresfordi (Fig. 5B).
Redescription:Male: Body length 6–7 mm.
Head: As in S. beresfordi except the following: ocellar triangle shiny and pale yellow, rarely light brown, surrounded by dull and darker V-shaped area (Fig. 4A); flagellum 2× length of the pedicel; scape appears subtly lighter than in S. beresfordi (S. beresfordi head, Fig. 2A).
Thorax: Indistinguishable from S. beresfordi (S. neglecta thorax, Figs. 4B, E, F).
Legs: Indistinguishable from S. beresfordi.
Wing: Costal vein covered in short setulae with one long black seta dorsal-basally.
Abdomen: Generally indistinguishable from S. beresfordi, although S. neglecta often tends to be a slightly lighter brown dorsally than S. beresfordi.
Terminalia; Slight lateral grove extends almost the entire length of more elongate surstylus (Fig. 5B); postgonite is somewhat more flattened, less scooping or cup posture than S. beresfordi (Fig. 5B).
Female: Similar to male except abdominal banding is somewhat lacking; abdomen terminates in long and slender ovipositor which closely resembles S. beresfordi (Fig. 2B); tergite 5 is about 5 mm long with anterior portion pale yellow, followed by darker brownish black, and abruptly terminating in paler yellow; majority of ovipositor with short, irregular setulae on dorsal surface only, becoming distinctly regular and forming thick band of black pile towards dark, anterior end of ovipositor, the pile becoming rather pale on distally palest area just before genitalia.
Terminalia: As in S. beresfordi except: cerci short, with very thick and long black spine-like setae, not extending to tip of hypoproct, setae sharply pointed (Fig. 4D).
Egg: Very similar to S. beresfordi. Only discernable difference is microsculptured surface (Fig. 4C) slightly more defined than S. beresfordi (Fig. 2D). Egg described in detail by Taber and Maloney (Reference Taber and Maloney2006).
Hosts: Unknown.
Known distribution:Stylogaster neglecta ranges from Ontario and southern Québec, Canada, south to Georgia and west as far as mid Nebraska and Kansas, United States of America (Fig. 6B). Camras and Parrillo (Reference Camras and Parrillo1985) suggested that the range extended farther west to Arizona and farther south to Sinaloa and Chiapas, Mexico. This is undoubtedly due to the inclusion of specimens of our new species, S. beresfordi, within the former boundaries of S. neglecta.
Comments: Five COI sequences have been obtained and are published in GenBank (Table 1). The sequences were identical. Genetically and morphologically, this species is closest to S. beresfordi and is hypothesised to be its sister clade (Fig. 7).
Acknowledgements
Thanks to Scott Kelso for collecting some of the COI barcoding data and to the Canadian Centre for DNA Barcoding for sequencing other specimens and housing the data in BOLD. Victoria Nowell databased and geocoded all of the specimens examined. This work was supported by funding to J.H.S. from Agriculture and Agri-Food Canada and the Natural Sciences and Engineering Research Council of Canada. David Moll provided the field photographs of S. beresfordi (Figs. 1B, C). Jeff Cumming provided valuable advice that helped develop the approach to the project. Jens-Hermann Stuke and one anonymous reviewer provided useful comments on the manuscript and helped to improve the final product. L.R. provided insight into Neotropical Stylogaster and examined type material of most world species. He also provided guidance with character selection. T.O.B. and J.H.S. worked together equally on specimen examination, species concept formulation, and data organisation. T.O.B. took the lead in preparation of the manuscript and key and produced all of the drawings used in the paper. J.H.S. reworked and fleshed out the manuscript.