


High dry-bulb temperature and humidity, in combination with a solar load and low air movement, can exceed stressor limits, resulting in reduced animal productivity and impaired animal well-being( Reference Silanikove 1 ). Baumgard & Rhoads( Reference Baumgard and Rhoads 2 ) have reported that heat stress markedly alters post-absorptive carbohydrate, lipid and protein metabolism independent of reduced feed intake through coordinated changes in fuel supply and utilisation by multiple tissues. Apart from its direct effects on nutrient intake and metabolism, much of the detrimental effects of heat stress on animal productivity may be indirectly mediated by reduced antioxidant capacity, inappropriate hormone secretions, and increased susceptibility to parasites and disease occurrence( Reference Mujahid, Pumford and Bottje 3 – Reference Rhoads, Kim and Collier 5 ).
In addition to functioning as the primary site of nutrient absorption, the gastrointestinal tract is also an important component of the immune system( Reference Baumgart and Dignass 6 – Reference Bailey, Haverson and Inman 8 ). The ambient temperature clearly affects the physiological functions of the gastrointestinal tract. Heat-stressed mammals channelise blood to the periphery of their body in an attempt to maximise radiant heat dissipation, and this blood redistribution is accompanied by vasoconstriction of the gastrointestinal tract( Reference Lambert 9 ). As a result, reduced blood and nutrient flow results in hypoxia at the intestinal epithelium, which ultimately compromises intestinal integrity and function( Reference Yan, Zhao and Wang 10 ). Consequently, heat-induced intestinal permeability is associated with the increased abundance of blood markers of endotoxaemia, hypoxia and inflammation, all of which may contribute to the multi-organ failure syndrome( Reference Hall, Buettner and Oberley 11 ). Damage to the mucosal epithelium of the small intestine caused by heat stress( Reference Liu, Yin and Du 12 ) can directly affect intestinal barrier function and nutrient absorption in the small intestine( Reference Boudry, Péron and Huërou-Luron 13 , Reference Lallès, Bosi and Smidt 14 ), resulting in reduced growth performance.
Management strategies are required to alleviate heat stress. Beede & Collier( Reference Beede and Collier 15 ) have identified three management strategies that minimise the effects of thermal stress: physical modification of the living environment; breeding of animals for heat tolerance; improvement of nutritional management. Limiting energy intake has been proven to effectively decrease heat generation( Reference Carstens, Johnson, Ellenberger, Van Der Honing and Close 16 ) and thereby decrease overall metabolic heat load in animals subjected to high ambient temperatures. In addition, dietary manipulation to enhance the antioxidant capacity of the body has been proven to be an important measure in the alleviation of heat stress. Certain trace minerals (such as cofactors of antioxidant enzymes such as Se in glutathione peroxidase (GSH-Px)) and several vitamins (such as vitamins E and C) are integral components of the antioxidant system( Reference Weiss, Hogan and Todhunter 17 ) and have heat stress-lowering effects( Reference Sejian, Singh and Sahoo 18 ).
In recent years, there has been growing interest in the use of grape seed extract (GSE) as a dietary antioxidant supplement( Reference Santos Buelgo and Scalbert 19 ). The antioxidant capacity of GSE is due to the presence of high proportions of phenolic compounds, primarily monomeric catechin and epicatechin, gallic acid, and oligomeric and polymeric procyanidins in grapes( Reference Arnous and Meyer 20 ). Previous studies have reported that GSE possesses strong antioxidant capacity in vitro – 20- and 50-fold greater than that of vitamins E and C, respectively( Reference Caillet, Salmiéri and Lacroix 21 – Reference Yilmaz and Toledo 23 ) – in addition to immunomodulatory functionality( Reference Katiyar 24 ). The present study was carried out to investigate the effects of GSE on the antioxidant capacity and immune status of jejunum epithelial cells (JEC) isolated from a steer subjected to heat stress treatment.
Materials and methods
The present experiment was conducted according to the Animal Care and Use Guidelines of the Southwest University, Chongqing, China.
Preparation of jejunum epithelial cells
The methods followed for JEC isolation were based on the procedures described by Sun et al. ( Reference Sun, Zhang and He 25 ). Briefly, the jejunum epithelia collected from a Simmental × Qinchuan steer (about 500 kg body weight) were washed with (1) rinsing solution 1 (5 × 105 IU/l of penicillin G sodium and streptomycin sulphate) and (2) rinsing solution 2 (250 μg/ml of gentamicin and 12·5 μg/ml of amphotericin B). Subsequently, the sample was minced into small pieces, placed in a 200 ml Erlenmeyer digestion flask containing 50 ml of a digestion solution (0·25 % trypsin and 0·02 % EDTA) and incubated in a low orbital, hot-air incubator at 37°C for 30 min. The digestion solution containing pieces of the tissue sample was mixed using a drip pipe for 3 min. After incubation, this mixture was transferred to a 50 ml centrifuge tube containing Dulbecco's modified Eagle's medium with 10 % fetal bovine serum (DMEM-FBS) and centrifuged at 600 g for 5 min at room temperature. The supernatant was collected and used for Ig assay. The precipitate containing JEC was washed four times with sterile Hanks' balanced salt solution with an antibiotic–antimycotic mixture (1 × 105 IU/l of penicillin G sodium, 1 × 105 IU/l of streptomycin sulphate and 0·25 mg/ml of amphotericin B as Fungizone).
Grape seed extract treatment and heat stress exposure
Based on the time difference in cell adhesion between the fibroblasts and JEC, the cells were cultured for 1·5 h in DMEM-FBS. The cells were then transferred to a twenty-four-microwell plate at a density of 1 × 106 cells/plate in DMEM-FBS and cultured for 24 h under standard conditions at 37°C in a 5 % CO2-humidified atmosphere. After 24 h of culturing, the plated cells were washed with DMEM-FBS and cultured for an additional 10 h in DMEM-FBS supplemented with GSE (Tarac Technologies) at a final concentration of 0, 10, 20, 40 or 80 μg/ml at 37°C. According to the manufacturer's product description, the GSE contained 90·1 % (w/w) polyphenols, 5·0 % (+)-catechin, 4·8 % ( − )-epicatechin, 2·4 % ( − )-epigallocatechin, 14·1 % dimeric proanthocyanidins, 11·6 % trimeric proanthocyanidins, 7·8 % tetrameric proanthocyanidins and 40·0 % polymeric proanthocyanidins. The doses of GSE added to the culture medium in the present study were determined by referring to the doses used in the studies of Kaur et al. ( Reference Kaur, Singh and Gu 26 ) and Roy et al. ( Reference Roy, Baliga and Elmets 27 ). The cultured plates were carefully sealed and placed in a water-jacketed incubator with humidified air mixed with 5 % CO2. There were forty-eight replicates for each level of GSE treatment, and a total of 240 replicates for five GSE concentrations were cultured at a time.
Following GSE-supplemented culturing, the cells were subjected to heat stress treatment. The forty-eight replicates of each level of GSE treatment were further cultured at temperatures of 37°C (normal temperature for cell culture), 40, 43 and 46°C for 2 h. Temperatures of 40, 43 and 46°C were selected to reflect temperatures potentially causing slight, harmful and serious or lethal heat stress to cells, respectively. The entire experiment consisted of five GSE concentrations crossed with four temperatures, forming a 5 × 4 factorial design with twelve replicates for each treatment. All the replicates were obtained on the same day.
After 2 h of incubation, four replicates in each treatment were removed and Alamar Blue solution was added to determine cell viability as described below. Additionally, four replicates were preserved at − 80°C after treatment with the TRIzol reagent (Invitrogen) for isolating RNA, and the remaining four replicates were used for various assays described below.
Alamar Blue assay for cell viability
Cell viability was measured according to the procedures described by Byth( Reference Byth, Mchunu and Dubery 28 ). Alamar Blue solution was added directly into the plate to a final concentration of 10 %, and the plate was further incubated at 37°C for 3 h. After the incubation period, fluorescence measurements were performed using a fluorescence reader (ELISA module, Anthos microplate spectrophotometer Zenyth 200; Anthos Labtec Instruments, GmbH) with the excitation/emission wavelengths set at 570/630 nm, the sensitivity gain at 50, and the temperature at 37°C. Cell viability (%) was calculated by the equation derived by Willard et al. ( Reference Willard, Merritt, Dean, Willard, Merrit, Dean and Settle 29 ).
Antioxidant enzyme activity assays
The superoxide dismutase (SOD), catalase (CAT) and GSH-Px activities, as well as malondialdehyde concentration, of JEC were determined using the appropriate assay kits (Jiancheng Bioengineering Institute) according to the manufacturer's instructions. The nitrite colouration method was used to determine SOD activity at an excitation wavelength of 550 nm by measuring the absorbance. CAT activity was measured at an excitation wavelength of 405 nm according to the method of Góth( Reference Góth 30 ) using an assay kit. GSH-Px activity was measured at an excitation wavelength of 412 nm by quantifying the rate of oxidation of reduced glutathione (GSH) to form oxidised GSH. The thiobarbital method was used to determine cellular malondialdehyde concentration at an excitation wavelength of 532 nm by measuring the absorbance( Reference Tang, Tian and Gao 31 ). The cellular protein concentration was determined using the Coomassie Brilliant Blue G250 reagent, with bovine serum albumin used as a standard.
Ig and cytokine assays
The concentrations of Ig were measured by solid-phase indirect ELISA as described by Islam et al. ( Reference Islam, Hossain and Kamal 32 ). The cellular concentrations of TNF-α, IL-1β, IL-8 and Toll-like receptor 4 (TLR4) were determined using bovine-specific commercial ELISA kits for TNF-α (E90133Bo; Uscn Life Science Inc.), IL-1β (E90563Bo; Uscn Life Science Inc.), IL-8 (E90080Bo; Uscn Life Science Inc.) and TLR4 (A90753Bo01; Uscn Life Science Inc.), respectively.
Total RNA was isolated from the cultured cells according to the manufacturer's instructions. First-strand complementary DNA was synthesised using the RevertAid™ First Strand cDNA Synthesis Kit (K1622; Fermentas Inc.). To quantify the mRNA of glyceraldehyde 3-phosphate dehydrogenase (GAPDH, a housekeeping gene)(
Reference Murthi, Fitzpatrick and Borg
33
), IL-1β, IL-8, TLR4, TNF-α, epidermal growth factor (EGF) and EGF receptor (EGFR), real-time PCR was carried out using the complementary DNA of JEC. Primer sequences for each gene are listed in Table 1. The specificity of the primers was tested with a BLAST analysis against the genomic NCBI database. Real-time PCR was carried out using the SYBR Green method and the ABI 7900 Sequence Detection System (Applied Biosystems). Analyses were carried out in triplicate, and mean values were calculated. Data were collected and analysed using the ‘fit point’ option of the LightCycler software (version 3.5; Idaho Technology Inc.). A calibration curve was generated by the amplification of serially diluted complementary DNA using the fit point option of the LightCycler software for the target genes and GAPDH gene as an internal reference. The threshold fluorescence level was determined within the geometric region of the semi-log view of the amplification plot. The relative expression of the target gene was calculated using the
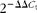 $$2^{ - \Delta \Delta C _{t}} $$
method(
Reference Livak and Schmittgen
34
).
$$2^{ - \Delta \Delta C _{t}} $$
method(
Reference Livak and Schmittgen
34
).
Table 1 Primer sequences for genes in the jejunum mucosa

GAPDH, glyceraldehyde 3-phosphate dehydrogenase; TLR4, Toll-like receptor 4; EGF, epidermal growth factor; EGFR, epidermal growth factor receptor.
Immunoblotting analysis
The cells were lysed on ice for 30 min in a radioimmunoprecipitation assay buffer (150 mm-NaCl, 50 mm-Tris–HCl, pH 8·0, 1 % Triton X-100, 1 % sodium deoxycholate and 0·1 % SDS) containing a protease inhibitor cocktail (1:1000 dilution; Sigma-Aldrich). Lysates were collected, passed through a 21G needle and centrifuged (14 000 g for 15 min at 4°C) to remove particulate matter. Protein content in the lysate supernatants was determined using DC Protein Assay reagents (Bio-Rad Laboratories) and bovine serum albumin as a standard. Equal quantities of protein (25 μg) were resolved using SDS–PAGE and transferred onto nitrocellulose membranes using semi-dry blotting techniques (Bio-Rad Laboratories). Non-specific protein-binding sites were blocked by treating the membranes with 5 % bovine serum albumin dissolved in Tris-buffered saline (10 mm-Tris–HCl, pH 8·0, and 150 mm-NaCl) with 0·05 % Tween-20. The membranes were incubated with primary anti-heat shock protein 70 (HSP70; no. 4825; Cell Signaling Technology, Inc.) and anti-GAPDH (no. GTX43800; Cell Signaling Technology, Inc.) antibodies, as well as horseradish peroxidase-conjugated secondary antibodies, in Tris-buffered saline with 0·05 % Tween-20 and 1 % bovine serum albumin. The bands were visualised using a chemiluminescence substrate (SuperSignal West Pico; Thermo Fisher Scientific) and fluorography. Densitometric analysis of the bands was carried out using an AlphaImager 2200 system (AlphaInnotech).
Statistical analyses
All statistical analyses were conducted using the generalised linear model procedure (SAS Institute, Inc.). The normality of data distribution was assessed using the Kolmogorov–Smirnov goodness-of-fit test. As the experiments were organised in a 5 × 4 factorial design, a two-way ANOVA was used to test the effects of GSE concentration, temperature and temperature × GSE concentration interactions, followed by a Tukey's post hoc test. Orthogonal contrasts were used to examine the linear and quadratic effects in response to increasing temperature or GSE concentration. All statistical tests were two-sided and statistical significance was set at P< 0·05.
Results
Supplementation with 20, 40 and 80 μg/ml GSE increased the GSH-Px activity of JEC cultured at 40°C (P= 0·04) and the CAT activity of JEC cultured at 43°C (P= 0·04; Table 2). On the whole, the activities of SOD (linear and quadratic, P< 0·001), CAT (linear and quadratic, P< 0·001) and GSH-Px (linear, P< 0·001; cubic, P= 0·02) of JEC declined in response to an increase in culture temperature. A rise in culture temperature increased the malondialdehyde concentration of JEC (linear and quadratic, P< 0·001; cubic, P= 0·01). Supplementation of the culture medium with GSE increased the activities of SOD (linear, P= 0·04), CAT (linear and quadratic, P< 0·001) and GSH-Px (linear, P= 0·01) of JEC.
Table 2 Effects of grape seed extract (GSE) and heat stress on the antioxidant capacity of jejunum epithelial cells of a Simmental×Qinchuan steer (Mean values and standard deviations; n 4)

SOD, superoxide dismutase; CAT, catalase; GSH-Px, glutathione peroxidase; MDA, malondialdehyde; T, temperature; T × GSE, interaction between T and GSE.
a,bMean values within a column with unlike superscript letters were significantly different (P <0·05).
When cultured at 40°C, JEC supplemented with 20, 40 and 80 μg/ml GSE exhibited IL-1β (P= 0·02) and TLR4 (P= 0·04) concentrations that were significantly higher than those in cells not supplemented with GSE (Table 3). When cultured at 43°C, JEC supplemented with 20, 40 and 80 μg/ml GSE exhibited TLR4 (P= 0·03) and TNF-α (P= 0·002) concentrations that were significantly lower than those in cells not supplemented with GSE. Overall, an increase was observed in the concentrations of IL-1β (linear and quadratic, P< 0·001; cubic, P= 0·003), IL-8 (linear and quadratic, P< 0·001; cubic, P= 0·02), TLR4 (linear and quadratic, P< 0·001) and TNF-α (linear, quadratic and cubic, P< 0·001) of JEC in response to an increase in culture temperature (P< 0·001). Supplementation of the culture medium with GSE decreased the concentrations of IL-1β (linear and quadratic, P< 0·001), IL-8 (linear and quadratic, P< 0·001), TLR4 (quadratic, P= 0·008; cubic, P= 0·002) and TNF-α (linear, P= 0·03) of JEC. Significant temperature × GSE interactions were observed for the concentrations of IL-1β (P< 0·001), IL-8 (P< 0·001) and TNF-α (P= 0·01) of cultured JEC.
Table 3 Effects of grape seed extract (GSE) and heat stress on the immune parameters of jejunum epithelial cells of a Simmental×Qinchuan steer (Mean values and standard deviations; n 4)

TLR4, Toll-like receptor 4; T, temperature; T × GSE, interaction between T and GSE.
a,b,cMean values within a column with unlike superscript letters were significantly different (P <0·05).
When cultured at 40°C, JEC supplemented with 40 and 80 μg/ml GSE exhibited decreased mRNA expression of IL-8 (P= 0·01) and TNF-α (P= 0·03) and increased mRNA expression of EGF (P= 0·03) when compared with those not supplemented with GSE (Table 4). When cultured at 43°C, JEC supplemented with 20, 40 and 80 μg/ml GSE exhibited mRNA expression of IL-8 (P< 0·001) and TLR4 (P= 0·002) that was lower than that in cells not supplemented with GSE. On the whole, increases were observed in the mRNA expression of IL-1β (linear, P< 0·001; quadratic and cubic, P= 0·002), IL-8, TLR4 and TNF-α (linear, quadratic and cubic, P< 0·001) of JEC in response to an increase in culture temperature and declines were observed in the mRNA expression of EGF (linear, quadratic and cubic, P< 0·001) and EGFR (linear and cubic, P< 0·001). Supplementation of the culture medium with GSE decreased the mRNA expression of IL-8 and TNF-α (linear, P= 0·002) and increased the mRNA expression of EGF and EGFR (linear, P< 0·001) of JEC.
Table 4 Effects of grape seed extract (GSE) and heat stress on the mRNA expression of cytokines of jejunum epithelial cells of a Simmental×Qinchuan steer (Mean values and standard deviations; n 4)

TLR4, Toll-like receptor 4; EGF, epidermal growth factor; EGFR, epidermal growth factor receptor; T, temperature; T × GSE, interaction between T and GSE.
a,b,cMean values within a column with unlike superscript letters were significantly different (P <0·05).
When cultured at 40°C, JEC supplemented with 40 and 80 μg/ml GSE exhibited viability that was greater than that of cells supplemented with 0 and 10 μg/ml GSE (P= 0·004) and those supplemented with 0 and 10 μg/ml GSE exhibited HSP70 concentration that was greater than that of cells supplemented with 40 and 80 μg/ml GSE (P< 0·001; Table 5). Similarly, when cultured at 43°C, JEC supplemented with 20, 40 and 80 μg/ml GSE exhibited viability that was greater than that of cells supplemented with 0 and 10 μg/ml GSE (P< 0·001) and those supplemented with 0 and 10 μg/ml GSE exhibited HSP70 concentration that was greater than that of cells supplemented with 20, 40 and 80 μg/ml (P< 0·001). Overall, the viability of JEC declined in response to an increase in culture temperature (linear and cubic, P< 0·001), whereas the concentration of HSP70 increased (linear, quadratic and cubic, P< 0·001). Supplementation of the culture medium with GSE increased the viability of JEC (linear, P< 0·001; quadratic, P= 0·002) and decreased the concentration of HSP70 (linear and quadratic, P< 0·001). Temperature × GSE interactions were detected for cell viability (P< 0·001) and HSP70 concentration of JEC (P= 0·004).
Table 5 Effects of grape seed extract (GSE) and heat stress on the cell viability and heat shock protein 70 (HSP70) concentration of jejunum epithelial cells of a Simmental×Qinchuan steer (Mean values and standard deviations; n 4)

T, Temperature; T × GSE, interaction between T and GSE.
a,bMean values within a column with unlike superscript letters were significantly different (P <0·05).
Discussion
The biological properties of polyphenols are dependent on the quantity of polyphenols consumed and on their bioavailability, which apparently differs greatly among the various polyphenols( Reference Manach, Williamson and Morand 35 ). Most polyphenols present in food exist in the form of esters, glycosides or polymers that cannot be absorbed in their native form. Typically, these substances must be cleaved before absorption( Reference Rios, Bennett and Lazarus 36 , Reference Serra, Macià and Romero 37 ). Polyphenols with relatively small molecular weights are easily absorbed through the gut( Reference Martin and Appel 38 ). Most of the absorbed polyphenols present in the circulation and tissues predominately exist in conjugated forms – glucuronised, sulphated or methylated – or in a combination of these forms( Reference Serra, Bryant and Motilva 39 ). We speculate that the bioavailability of polyphenols in the GSE used in the present study would be high because it contains a relatively high proportion of monomeric or oligomeric polyphenols. However, data concerning differences in bioavailability between GSE polyphenols and dietary polyphenols are limited and therefore further study is required.
Antioxidant capacity
The antioxidation defence network of animal organs and tissues acting against oxygen-free radicals includes endogenous (e.g. SOD, GSH, GSH-Px and CAT) and exogenous (e.g. vitamin E and Se) factors( Reference Fang, Yang and Wu 40 ). In the present study, the activities of SOD, CAT and GSH-Px of JEC declined in response to an increase in culture temperature, whereas the cellular malondialdehyde concentration increased. These results indicate that heat stress decreases the antioxidant capacity of JEC and are in agreement with previous findings. Burke et al. ( Reference Burke, Scaglia and Saker 41 ) reported that heat stress decreases the activities of GSH reductase and GSH-Px of peripheral blood mononuclear cells of heifers. Heat stress has been found to enhance the formation of reactive oxygen species and disturb the balance between the production of free radicals and the antioxidant systems, ultimately leading to oxidative stress( Reference Lord-Fontaine and Averill-Bates 42 – Reference Sahin, Onderci and Sahin 46 ). Mujahid et al. ( Reference Mujahid, Yoshiki and Akiba 47 ) also reported that heat stress increases the generation of free radicals, possibly through the disruption of electron transport assemblies of the mitochondrial membrane. Moreover, heat stress has been shown to decrease intestinal blood flow and ultimately reduce the absorption of nutrients, including exogenous antioxidant nutrients( Reference Payne and Southern 48 ).
In the present study, the CAT activity of JEC cultured at 43°C and the GSH-Px activity of those cultured at 40°C increased with GSE supplementation. This result indicates that GSE can assist in the maintenance of redox balance in JEC cultured at 40 and 43°C. Primarily notable for the bioactivity of its phenolic compounds, GSE and its antioxidative characteristics– including scavenging of free radicals, inhibition of lipid oxidation and reduction of hydroperoxide formation– have been widely investigated( Reference Meyer, Yi and Pearson 49 , Reference Sato, Ramarathnam and Suzuki 50 ). The highest antioxidant capacity in grapes has been found in seeds, followed by skin, whereas the flesh has been found to have the lowest antioxidant capacity( Reference Pastrana-Bonilla, Akoh and Sellappan 51 ). Some clinical data have shown that procyanidin oligomers present in grape seeds are twenty times more potent than vitamin C and fifty times more potent than vitamin E as antioxidants( Reference Uchida, Edamatsu and Hiramatsu 22 ). The antioxidant activity of GSE has also been reported to improve the oxidative stability of a variety of biological samples – including cooked beef( Reference Ahn, Jeon and Lee 52 ), rats( Reference Chis, Ungureanu and Marton 53 ), turkey, and pork patties, and cold-stored turkey meat( Reference Carpenter, O'Grady and O'Callaghan 54 – Reference Mielnik, Aaby and Skrede 56 ) – based on in vitro and in vivo studies. Many investigators have attempted to identify the phenolic compounds and chemical structure(s) that are primarily responsible for the antioxidant activity of grape extracts. Faria et al. ( Reference Faria, Mateus and de Freitas 57 ) observed that in five fractions with differing compositions of procyanidin polymers, the dimers displayed the highest antioxidant capacity based on a molar base (scavenging peroxyl radicals). A similar result was observed by Soobrattee et al. ( Reference Soobrattee, Neergheena and Luximon-Rammaa 58 ), who reported that the compound with the highest antioxidative activity among the variety of phenolic compounds was the procyanidin dimer and that antioxidant capacity decreased in the order of procyanidin dimer>flavanol>flavonol>hydroxycinnamic acids>simple phenolic acids. Diphenols are more effective antioxidants than simpler phenols due to the stabilisation of the phenoxy radical through hydrogen bonding( Reference Amico, Chillemi and Mangiafico 59 ). The high-molecular-weight compounds may be functionally as important as the monomer flavanols, such as catechin, which have been found to display high antioxidant potential among the phenolic compounds( Reference Yilmaz and Toledo 23 ). Furthermore, the antioxidant activity of a sample could be a function of a synergic effect among several components rather than of a single compound( Reference Maier, Schieber and Kammerer 60 , Reference Monagas, Hernández-Ledesma and Garrido 61 ).
Immunity
In the present study, the protein concentrations of IgG, IL-1β, IL-8, TLR4 and TNF-α and the mRNA expression levels of IL-1β, IL-8, TLR4 and TNF-α in JEC increased in response to an increase in culture temperature. As an important component of the immune system( Reference Baumgart and Dignass 6 – Reference Bailey, Haverson and Inman 8 ), the gastrointestinal tract initiates protective immune responses induced upon encountering pathogens and toxins or tolerance to commensal bacteria and food antigens. Under normal health conditions, T cells appear to be activated but unstimulated (secreting cytokines), but are prepared for stimulation and to secrete a variety of cytokines, including IL-2, IL-4, IL-5, IL-10 and interferon-γ( Reference Braunstein, Qiao and Autschbach 62 ). Once invaded, the intestinal epithelium can function as a component of the innate immune system by expressing TLR (e.g. TLR2, TLR3, TLR4 and TLR5) and secrete cytokines (e.g. TNF-α, IL-15 and IL-8)( Reference Cario and Podolsky 63 – Reference Maaser and Kagnoff 66 ). High circulating concentrations of cytokines in the plasma – including IL-1β, IL-6 and IL-10 – have also been observed in heat stroke-affected animals and humans( Reference Bouchama, Roberts and Al Mohanna 67 , Reference Leon, Blaha and DuBose 68 ). Heat shock has been shown to induce (in cultured intestinal epithelial cells) the production of cytokines (e.g. IL-6 and IL-8)( Reference Wang, Guo and Noel 69 ), directly resulting in increased epithelial permeability and a high rate of apoptosis( Reference Hu, Claud and Musch 70 ). Such a heightened immune response could result in a systemic inflammatory reaction, ultimately leading to multiple-organ failure, such as that observed during sepsis( Reference Lambert 9 ).
In the present study, GSE supplementation was found to decrease the protein concentrations of IgG, IL-1β, IL-8, TLR4 and TNF-α and the mRNA expression levels of IL-8, TNF-α and TLR4 in JEC. This result indicates that GSE assists in the suppression of inflammatory reaction of JEC subjected to heat stress treatment. The immunoregulatory function of GSE can be attributed to its chemical composition. Phenolic compounds in grape seeds have been shown to exert significant anti-inflammatory effects in rats, mice and humans( Reference Chacóna, Ceperuelo-Mallafré and Maymó-Masipa 71 , Reference Panico, Cardile and Avondo 72 ), with the contributing molecules purported to include flavonols, flavanols and procyanidins (oligomeric flavonoids)( Reference Chacóna, Ceperuelo-Mallafré and Maymó-Masipa 71 , Reference Terra, Montagut and Bustos 73 ). Bralley et al. ( Reference Bralley, Hargrove and Greenspan 74 ) reported that ear inflammation, oedema and polymorphonuclear leucocyte infiltration induced by 12-O-tetradecanoylphorbol 13-acetate were inhibited in mice treated for 30 min with extracts from grape skins and seeds. These findings indicate that phenolic compounds in grapes apparently possess anti-inflammatory activity. The mechanisms responsible for the anti-inflammatory activity of procyanidins have also been investigated in previous studies. Inhibition or reduction of cytokine gene expression may be an important pathway involved in the anti-inflammatory activity of grape phenolic compounds( Reference Chacóna, Ceperuelo-Mallafré and Maymó-Masipa 71 , Reference Terra, Montagut and Bustos 73 ). Human adipocytes and macrophage-like cell lines have been found to produce low amounts of IL-6 and monocyte chemotactic protein-1 and high amounts of anti-inflammatory adipokine and adiponectin after pre-treatment with extracts of grape seed procyanidins( Reference Chacóna, Ceperuelo-Mallafré and Maymó-Masipa 71 ). These results demonstrate that the regulation of the expression of anti-inflammatory and pro-inflammatory cytokines is the primary mechanism responsible for the roles of GSE in the immunoregulatory function of JEC subjected to heat stress treatment.
Epidermal growth factor and epidermal growth factor receptor
In the present study, the mRNA expression levels of EGF and EGFR in JEC decreased with an increase in culture temperature. Liu et al. ( Reference Liu, Yin and Du 12 ) also observed that under high ambient temperatures, a treatment regimen of 40°C for 5 h per d over a 10 d period decreased the mRNA expression levels of EGF and EGFR in porcine small-intestinal epithelia. Previous reports indicate that the expression of EGF and EGFR is related to the damage and recovery of small-intestinal mucosa( Reference Helmrath, Shin and Erwin 75 , Reference Nair, Warner and Warner 76 ). Many investigators have considered that EGF may have no effect on normal intestinal mucosal epithelium, but rather may promote cell proliferation, repair and migration during regeneration following damage( Reference Helmrath, Shin and Erwin 75 , Reference Nair, Warner and Warner 76 ). The signalling cascade mediated by EGFR is characterised by pleiotropy, including cell proliferation, repair and migration, and internal environment stabilisation( Reference Helmrath, Shin and Erwin 75 ). Therefore, we speculate that the cell proliferation of JEC subjected to heat stress treatment is likely to be decreased, accompanied by a decline in EGF/EGFR gene expression, a hypothesis supported by the observation that the viability of JEC declined in response to an increase in culture temperature.
In the present study, we found that GSE supplementation increased the mRNA expression levels of EGF and EGFR in JEC cultured at 40 or 43°C. This result indicates that GSE can enhance the growth of normal cells through the activation of EGF-induced signalling pathways. Previous studies have also reported that GSE exhibits the beneficial property of enhancing the growth and viability of normal cells( Reference Bagchi, Bagchi and Stohs 77 ) while exhibiting cytotoxicity towards some types of cancer cells( Reference Sharma, Tyagi and Singh 78 , Reference Meeran and Katiyar 79 ). However, the mechanism by which GSE can distinguish tumour cells from normal cells remains unclear.
Heat shock protein 70
In present study, the protein concentration of HSP70 in JEC increased with elevated culture temperatures. HSP70 is the most investigated member of a family of proteins synthesised in response to physical, chemical or biological stresses, including heat exposure( Reference Deane and Woo 80 – Reference Staib, Quindry and French 82 ). The expression of HSP70 is rapidly up-regulated under conditions of oxidative stress, providing cytoprotection by making cells resistant to otherwise lethal levels of environmental stress( Reference Kampinga 83 ). There have been many published reports about the protective effects of HSP70 on the intestine under conditions of heat or other stresses in both in vitro and in vivo models( Reference Wischmeyer, Musch and Madonna 84 – Reference Ohkawara, Nishihira and Takeda 86 ). HSP70 develops a protective interaction with sarco(endo)plasmic reticulum Ca2+-ATPases 2a (SERCA2a) during heat stress, reducing the oxidation and nitrosylation of SERCA2a and thereby increasing its maximal activity( Reference Fu and Tupling 87 ).
In the present study, GSE supplementation was found to decrease the HSP70 concentration of JEC. Kim et al. ( Reference Kim, Deshane and Barnes 88 ) also reported that the proteins affected by GSE included those involved in energy generation and protein folding (e.g. HSP60, HSP70 and HSP71, cytoskeletal proteins, and medium-chain and glial fibrillary acidic protein). According to antioxidant capacity and immunity analyses, we speculate that the decline in the expression of HSP70 in heat-induced JEC supplemented with GSE may be closely related to the increase in antioxidant capacity and the decline in inflammatory reactions.
Cell viability
In the present study, heat stress-induced cytotoxicity was assessed using a cell viability assay, which confirmed that heat stress decreased cell viability. Heat stress has also reportedly been found to decrease cell viability and induce cell death in bovine mammary epithelial cells( Reference Du, Di and Guo 89 ) and hepatic cells of grass carp( Reference Cui, Liu and Xie 90 ). Heat stress and heat stress-induced oxidative stress have been found to adversely affect the structure and physiology of the cell (e.g. impairing transcription, RNA processing and translation and altering membrane structure)( Reference Iwagami 91 , Reference Sahin, Orhan and Akdemir 92 ) and decrease cell viability( Reference Du, Di and Guo 89 , Reference Cui, Liu and Xie 90 ). Based on the results of the present study, we speculate that the decreased cell viability of JEC subjected to heat stress treatment may be highly related to changes in antioxidant capacity, inflammatory reaction, HSP70 expression, etc.; however, the underlying mechanisms remain unclear and require further study.
In the present study, GSE supplementation was also found to increase the cell viability of JEC cultured at 40 or 43°C. Similarly, cocoa phenolic extract has been found to exhibit a remarkable recovery in cell viability after damage induced by tert-butylhydroperoxide( Reference Martín, Ramos and Cordero-Herrero 93 ). However, previous studies have also shown that phenols can significantly impair the viability of human colonic epithelial cells( Reference Pedersen and Saermark 94 ). These phenol-induced effects are secondary to changes occurring in the cell membrane resulting in the disruption of tight junction-containing microdomains, and the disruption of these microdomains results in the mislocalisation of tight junction components and altered paracellular permeability( Reference McCall, Betanzos and Weber 95 ). In general, previous data on cell viability of humans or animals exposed to plant phenols are inadequate to establish dose–response relationships and further study is required to establish these relationships.
Conclusion
We report that heat stress decreases the antioxidant capacity of JEC in steers and activates the immune response by up-regulating the expression of anti-inflammatory cytokines in cells. When cultured at 40 and 43°C, but not at 37 and 46°C, GSE attenuated the heat stress-induced responses of small-intestinal epithelial cells obtained from a cross-bred steer by increasing the antioxidant capacity and suppressing the immune response. Taken together, we conclude that GSE attenuates the responses of JEC of Simmental × Qinchuan steers to heat stress, primarily by increasing antioxidant capacity and cell viability and decreasing inflammatory reactions. However, GSE played no role in the regulation of the responses of JEC cultured at normal or extremely high temperatures. Furthermore, a dose–response in vivo study with GSE (based on the doses evaluated in the present study) is required to obtain information regarding the practical physiological and biochemical effects of GSE supplementation, as well as the lowest effective and the highest non-toxic doses of GSE necessary for optimum heat stress studies.
Acknowledgements
The present study was financially supported by the National Science & Technology Pillar Program (2012BAD14B18, 2014BAD08B11 and 2011BAD36B01), which contributed to the study design, study conduct, analysis of the samples and data, and interpretation of the findings, and the National Program on Key Basic Research Project of China (2013CB127303 and 2013CB127304), the Natural Science Foundation Project of CQ CSTC (cstc2013jjB0112 and cstc2012jjA80001), the Fundamental Research Funds for the Central Universities (XDJK2011C030 and XDJK2011B011) and the National Natural Science Foundation of China (30600436), which contributed to the analysis of the samples and data, interpretation of the findings and preparation of the manuscript.
The authors' contributions are as follows: Z. S. and X. L. wrote the initial draft of the manuscript; X. L., Y. Y., C. C., J. Y. and Z. S. implemented and conducted the study, collected the data and analysed the data; Z. S. and S. L. interpreted the data and provided critical comments; S. L. revised the manuscript.
None of the authors has any conflicts of interest to declare.





