


Osteoporosis is a metabolic condition characterised by low bone mass, deterioration of bone tissues and increased risk of fracture(Reference Cooper, Westlake and Harvey1). It is a worldwide public health problem that creates a significant economic burden on society as well as to the families of patients who suffer from related fractures(Reference Cooper, Westlake and Harvey1, Reference Dennison, Cole and Cooper2). Hormone replacement therapy used to be the major regimen for the prevention and treatment of postmenopausal osteoporosis. However, with the recent discovery of the association between hormone replacement therapy and the increased risks in postmenopausal women to develop breast, endometrial or ovarian cancers(Reference Zhang, Chen and Lai3–Reference Palacios5), there is a strong demand to develop alternative approaches for the management of osteoporosis.
Isoflavones are compounds structurally similar to the mammalian oestrogens that are found in plant foods, particularly soyabeans. They have received considerable attention for their preventive actions on bone loss(Reference Hertrampf, Gruca and Seibel6). Genistein (4′,5,7-trihydroxyisoflavone) is the major isoflavone found in these foods(Reference Szkudelska and Nogowski7). The potential efficacy of genistein on bone biology has been extensively studied and it mainly acts as a selective oestrogen receptor modulator in vivo and in vitro to exert beneficial effects on bones(Reference Zhang, Chen and Lai3).
Erythrina variegata L. (EV; Leguminosae), which has been used as folk medicine in the tropical and subtropical regions, is known to possess pharmacological activities including antimicrobial(Reference Sato, Tanaka and Yamaguchi8), anti-inflammatory(Reference Cox9) and antibacterial effects(Reference Sato, Tanaka and Fujiwara10). Phytochemical work has demonstrated that EV is rich in genistein derivatives(Reference Zhang, Li and Yao11, Reference Li, Wang and Wong12), which is a possible explanation for the efficacy of using EV extract for the prevention and treatment of ovariectomy-induced osteoporosis in rats as reported in our previous study(Reference Zhang, Li and Lai13). However, the mechanisms by which EV exerts its protective actions on bone have yet to be studied.
In an attempt to explore the potential mechanism involved in the osteoprotective effects of EV, the dose-dependent effects of EV on bone micro-structure and morphology as well as tibial bone-specific gene expression in ovariectomised rats, a widely used and well-established animal model for the study of postmenopausal osteoporosis, was used. In addition, the in vitro effects of EV on osteoclast differentiation in murine macrophage RAW 264·7 cells, which are recognised as an osteoclast precursor cell line, were also studied.
Materials and methods
Plant material and extraction
The collection and authentication of EV and EV extracts were done as previously described(Reference Zhang, Li and Lai13).
Animal study
Sixty virgin female Sprague–Dawley rats, aged 4 months, were purchased from Guangzhou University of Traditional Chinese Medicine (Guangzhou, China). Husbandry of the animals was carried out as previously described(Reference Zhang, Li and Lai13). All animals were allowed free access to distilled water and pair-fed a phyto-oestrogen-low diet (D00031602; Research Diets Inc., New Brunswick, NJ, USA), which was prepared according to the American Institute of Nutrition (AIN)-93M formulation (D10012M; Research Diets Inc.) except that maize oil was used instead of soyabean oil. The formulations and estimated nutrient composition of the experimental diets are referred to in our previously published paper(Reference Zhang, Li and Wan14). The acclimatised rats underwent either bilateral laparotomy (sham; n 10) or bilateral ovariectomy (n 50). At 1 week after healing from surgical procedures, the ovariectomised rats were randomly divided into five groups of ten: vehicle-treated, ovariectomy only (OVX), genistein-treated (40 mg/kg), low dose of EV extract-treated (200 mg/kg), medium dose of EV extract-treated (500 mg/kg) and high dose of EV extract-treated (1000 mg/kg). The EV extract and genistein were administered daily through a gastric tube. The dosages of EV used in the present study were chosen based on the maximal tolerated dose of EV determined in our previous study(Reference Zhang, Li and Lai13). The animals were on the diet and treatments for 12 weeks. At the end of the treatment, a 24 h urine sample was collected. The animals were then killed by withdrawing blood from the abdominal aorta under light diethyl ether anaesthesia. Serum was then prepared, and tibiae were dissected from soft tissue and stored frozen until used for RNA analysis. The animal study protocols were approved by the Animal Ethics Committee of The Hong Kong Polytechnic University.
Chemistry of serum and urine
Serum Ca, P and alkaline phosphatase (ALP), and urine Ca, P and creatinine were measured as previously described(Reference Zhang, Lai and Leung15).
Analysis of the micro-computed tomography scans
The tibia was scanned with a micro vivaCT 40 system (Scanco Medical, Bassersdorf, Switzerland). Thirty slices of captured images were established as the volume of interest. The trabecular bone was separated from the cortical bone by free drawing regions of interest using the software provided with the scanner. Morphologic measurements for the thirty slices were performed and the following three-dimensional parameters were obtained for the trabecular bone: (1) trabecular number; (2) trabecular thickness; (3) trabecular separation; (4) connectivity density (Conn.D). The following diameters were obtained for the cortical bone: (1) bone surface/bone volume (BS/BV); (2) Conn. D; (3) structure model index (SMI), which is a parameter used for the quantification of the ‘rod-like’ or ‘plate-like’ architecture. The SMI values for ideal plates and rods are 0 and 3, respectively.
RNA isolation and real-time RT-PCR
The extraction of total RNA from the tibia was performed according to the TRIzol manufacturer's protocol. First-strand cDNA were synthesised from 4 μg of total RNA in 20 μl reaction volume using the SuperScript First-Strand Synthesis System (Invitrogen, Carlsbad, CA, USA) and oligo dT random primers. Real-time PCR was performed in the LightCycler system as described(Reference Zhang, Lai and Leung16). The relative mRNA amount was normalised to glyceraldehyde 3-phosphate dehydrogenase mRNA. The PCR primers used in the present study were similar to those reported in our previous study(Reference Zhang, Dong and Leung17).
Cell culture
The murine macrophage cell line RAW 264·7 was maintained in phenol red-free Dulbecco's modified Eagle's medium containing 5 % charcoal-stripped fetal bovine serum (GIBCO-BRL, Gaithersburg, MD, USA) and antibiotics at 37°C under 5 % CO2. Cells (1 × 105) were seeded in a six-well plate and cultured with receptor activator of nuclear factor-κB ligand (RANKL) (100 ng/ml) for osteoclast differentiation, simultaneously supplemented with vehicle or EV extract at the concentration of 10 μg/ml and 100 μg/ml in Dulbecco's modified Eagle's medium for 4 d. The concentration of EV was chosen based on the experimental results reported by Li(Reference Li18), who demonstrated that the effective dose range of EV in rat osteoblast-like cells was from 2 to about 200 μg/ml. The new medium was changed every 2 d.
Tartrate-resistant acid phosphatase staining
Tartrate-resistant acid phosphatase (TRAP) staining was carried out with a TRAP kit (Sigma, St Louis, MO, USA) according to the manufacturer's instructions. Briefly, cells were washed with PBS and treated with a fixation solution at room temperature for 5 min. They were then washed with distilled water and treated with a TRAP reagent at 37°C for 20–60 min. After washing with distilled water, the cells were observed under a microscope. Cells that possessed three or more nuclei were identified as multinucleated cells. TRAP-positive and multinucleated cells were counted as osteoclast-like cells.
Statistical analysis
The data from these experiments were reported as mean values with their standard errors for each group. All statistical analyses were performed using PRISM version 4.0 (GraphPad, San Diego, CA, USA). Inter-group differences were analysed by one-way ANOVA, and followed by Tukey's multiple comparison test as a post hoc test to compare the group means if overall P < 0·05. Differences in P < 0·05 were considered statistically significant.
Results
Weight gain, and serum and urine chemistry
As shown in Table 1, the weight gain increase in the OVX rats was more significant than in the sham rats (P < 0·05) despite the pair-feeding of the animals. Treatment of the ovariectomised rats with either the high dose of the EV extract or genistein reduced the ovariectomy-induced increase in weight gain (v. OVX rats, P < 0·05). Ovariectomy significantly decreased serum Ca (P < 0·01) and P (P < 0·05) levels and increased urinary Ca (P < 0·01) and P (P < 0·05). In addition, serum ALP levels increased in response to ovariectomy in the rats (P < 0·01). Treatment with genistein increased serum Ca (P < 0·01) and suppressed urinary Ca (P < 0·05) and P (P < 0·05) excretion as well as serum ALP levels (P < 0·01) in the ovariectomised rats. Similarly, treatment with the high dose of EV increased serum Ca (P < 0·05), suppressed urinary Ca (P < 0·05), P (P < 0·01) excretion and serum ALP levels (P < 0·01) in the ovariectomised rats. The two lower doses of EV had no significant effect on serum or urine chemistry as compared with ovariectomy alone.
Table 1 Effects of Erythrina variegata L. (EV) on weight gain and levels of biochemical markers in serum and urine of ovariectomised rats
(Mean values with their standard errors for nine or ten rats per group)
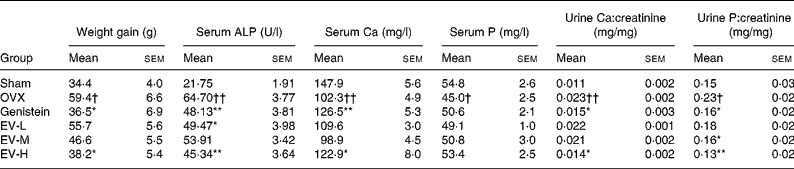
ALP, alkaline phosphatase; OVX, ovariectomised only (vehicle-treated); EV-L, low dose of EV extract; EV-M, medium dose of EV extract; EV-H, high dose of EV extract.
Mean value was significantly different from that of the OVX group: * P < 0·05, ** P < 0·01.
Mean value was significantly different from that of the sham-operated group: †P < 0·05, ††P < 0·01.
Micro-computed tomography analysis of trabecular bone and cortical bone
Fig. 1 summarises the result of micro-computed tomography analysis of the tibial trabecular bone and cortical bone in rats. For the trabecular bone of the tibial head, the trabecular number, trabecular thickness and Conn.D were significantly lower in the OVX group than in the sham group (Fig. 1(a), (b) and (d); P < 0·05), and trabecular separation was much higher in the OVX group than in the sham group (Fig. 1(c); P < 0·01). Administration of the high dose of EV (1000 mg/kg) increased trabecular number and Conn.D of the trabecular bone at the tibial head in the ovariectomised rats (P < 0·05), while genistein treatment did not lead to any improvement in these parameters of the trabecular bone. In addition, ovariectomy resulted in a dramatic increase in BS/BV and SMI and a reduction of Conn.D of the cortical bone of the tibial diaphysis in the rats. Treatment with genistein significantly suppressed the induction of SMI (P < 0·015) as well as the reduction of Conn.D (P < 0·01) in the ovariectomised rats. Treatment with the medium dose of EV suppressed the ovariectomy-induced increase in SMI level (P < 0·01) in the ovariectomised rats. Treatment of the ovariectomised rats with the high dose of EV significantly attenuated the changes in the cortical bone parameters (BS/BV, SMI and Conn.D) induced by ovariectomy (v. OVX, P < 0·05).

Fig. 1 Micro-computed tomography bone parameters of trabecular bone (a–d) and cortical bone (e–g) of tibial proximal metaphysis and diaphysis, respectively, in sham-operated, ovariectomised only (vehicle-treated) (OVX), and genistein- (Gen) or Erythrina variegata L. (EV)-treated ovariectomised rats. EV-L, low dose of EV extract; EV-M, medium dose of EV extract; EV-H, high dose of EV extract; Tb.N, trabecular number; Tb.Th, trabecular thickness; Tb.Sp, trabecular separation; Conn.D, connectivity density; BS/BV, bone surface/bone volume; SMI, structure model index. Values are means (n 10 per group), with standard errors represented by vertical bars. Mean value was significantly different from that of the OVX group: * P < 0·05, ** P < 0·01. Mean value was significantly different from that of the sham-operated group: † P < 0·05, †† P < 0·01.
Bone-specific gene expressions
Core-binding factor α1 (Cbfa1) is an osteoblast-specific transcription factor that plays an essential role in osteoblastic differentiation and maturation(Reference Zhang, Dong and Leung17), while cathepsin K (CtsK) is an osteoclast-specific protein that plays an important role in the collagen degradation phase of bone resorption(Reference Hie, Shimono and Fujii19, Reference Pennypacker, Shea and Liu20). As expected, the expression of Cbfa1 mRNA was suppressed (Fig. 2(a); P < 0·05) and the expression of CtsK mRNA was induced (Fig. 2(b); P < 0·05) in rat tibia in response to ovariectomy. Treatment of the ovariectomised rats with genistein reversed the changes in Cbfa1 and CtsK mRNA in tibia induced by ovariectomy (P < 0·01). In contrast, treatment of the ovariectomised rats with EV extract did not alter Cbfa1 mRNA expression in tibia, suggesting that EV extract might not alter bone formation in vivo. Furthermore, treatment of the ovariectomised rats with the medium or high dose of EV significantly down-regulated CtsK mRNA expression in tibia, suggesting that EV extract might suppress bone resorption in vivo (P < 0·05).

Fig. 2 Expressions of core-binding factor α1 (Cbfa1; a) and cathepsin K (CtsK; b) mRNA in the tibia of sham-operated, ovariectomised only (vehicle-treated) (OVX), and genistein- (Gen) or Erythrina variegata L. (EV)-treated ovariectomised rats. EV-L, low dose of EV extract; EV-M, medium dose of EV extract; EV-H, high dose of EV extract; GAPDH, glyceraldehyde 3-phosphate dehydrogenase. Values are means (n 10 per group), with standard errors represented by vertical bars. Mean value was significantly different from that of the OVX group: * P < 0·05, ** P < 0·01. † Mean value was significantly different from that of the sham-operated group (P < 0·05).
Osteoprotegerin (OPG) and RANKL, which are produced by osteoblasts, play an essential role in controlling the process of osteoclastogenesis(Reference Zhang, Lai and Leung16). mRNA expression of OPG (Fig. 3(a); P < 0·01) as well as the ratio of OPG:RANKL mRNA expression (Fig. 3(c); P < 0·05) in tibia decreased in response to ovariectomy as compared with the sham rats. Treatment of the ovariectomised rats with genistein (P < 0·05) or EV extracts at all three doses (P < 0·01) induced the expression of OPG in tibia, resulting in an increase in the OPG:RANKL ratio (P < 0·05) back to the sham level. However, RANKL mRNA expressions were not altered by ovariectomy or treatment with either genistein or EV extract.

Fig. 3 Expressions of osteoprotegerin (OPG; a) mRNA and receptor activator of nuclear factor-κB ligand (RANKL; b) mRNA and the OPG:RANKL ratio (c) in the tibia of sham-operated, ovariectomised only (vehicle-treated) (OVX), and genistein- (Gen) or Erythrina variegata L. (EV)-treated ovariectomised rats. EV-L, low dose of EV extract; EV-M, medium dose of EV extract; EV-H, high dose of EV extract; GAPDH, glyceraldehyde 3-phosphate dehydrogenase. Values are means (n 10 per group), with standard errors represented by vertical bars. Mean value was significantly different from that of the OVX group: * P < 0·05, ** P < 0·01. Mean value was significantly different from that of the sham-operated group: † P < 0·05, †† P < 0·01.
Tartrate-resistant acid phosphatase-positive cells
To determine if EV extract can directly regulate the process of osteoclast formation, pre-osteoclastic RAW 264·7 cells stimulated with RANKL were co-treated with the vehicle, or EV extract at 10 μg/ml or 100 μg/ml for 4 d. The present results indicated that TRAP-positive and multinucleated cells that possessed three or more nuclei were formed in RAW 264·7 cells in response to the RANKL stimulation, thus confirming the formation of osteoclasts (Fig. 4). EV at 100 μg/ml, but not at 10 μg/ml, dramatically reduced the number of TRAP-positive cells (P < 0·05) when compared with the vehicle-treated cells. These results indicated that EV at high concentrations could directly suppress the process of osteoclast formation in vitro.

Fig. 4 Tartrate-resistant acid phosphatase-positive (TRAP+) cell numbers in receptor activator of nuclear factor-κB ligand (RANKL) (100 ng/ml)-induced RAW 264·7 cells in the absence (control; Con) or presence of Erythrina variegata L. (EV) extracts at 10 μg/ml and 100 μg/ml for 4 d. Three separate experiments were performed with four repeated wells for each group. MNC, multinucleated cells; → ,TRAP+ MNC. Values are means with their standard errors. * Mean value was significantly different from that of the control treatment (P < 0·05).
Discussion
The present study demonstrated that ovariectomy induced higher weight gain, increases in serum ALP levels, urinary Ca and P excretion, and decreases in serum Ca and P levels, suggesting that ovariectomy led to changes in energy metabolism as well as bone and mineral metabolism. These changes are consistent with the results of our previous animal studies(Reference Zhang, Li and Lai13, Reference Zhang, Lai and Leung15) as well as other clinical studies(Reference Odvina, Poindexter and Peterson21, Reference Sahota, Mundey and San22). The ovariectomy-induced changes of biochemistry were simultaneously accompanied with bone loss and bone micro-architectural deterioration in both the trabecular bone and cortical bone. All these pathological alterations, including alterations in chemical markers and bone properties, demonstrated the onset and development of postmenopausal osteoporosis induced by oestrogen deficiency.
In the present study, EV extract could effectively inhibit the high rate of urinary Ca and P excretion in the ovariectomised rats and restore serum Ca and P levels to the normal level. Moreover, EV extract suppressed high bone turnover rate in the ovariectomised rats as indicated by a reduction in serum ALP level in the EV-treated groups as compared with that of the OVX rats. Therefore, EV might protect oestrogen-deficient animals from postmenopausal bone loss by reducing urinary mineral excretion, maintaining serum mineral levels as well as normal rate of bone turnover.
The results indicated that the micro-architecture of the trabecular bone at the tibial metaphysis was improved in the ovariectomised rats in response to the 12-week treatment of orally administrated EV extracts. These results were in agreement with the histomorphometric analysis performed on the same bone site in our previously reported study(Reference Zhang, Li and Lai13). In addition, EV extract decreased the value of SMI at the tibial diaphysis in the ovariectomised rats in a dose-dependent manner. Thus, EV helped to prevent the ovariectomy-induced formation of rod-like structures in the cortical bone of rat tibia. Furthermore, the cortical bone parameters of the ovariectomised rats treated with the high dose of EV extract, such as BS/BV and Conn.D, were restored almost to the level found in the tibial cortical bone of the sham-operated rats. These results suggest that EV extract could improve bone structure and morphology at both the trabecular and cortical bones in the ovariectomised rats. The improvement that EV brought to the bone structure and morphology of the tibial mid-shaft might account for the increase in bone biomechanical strength observed in our earlier report(Reference Zhang, Li and Lai13).
Genistein (40 mg/kg) did not display any beneficial effects against the ovariectomy-induced deterioration of the trabecular bone of rats in the present study. Similar results had been observed and reported in our recently published paper(Reference Zhang, Li and Wan14) in which genistein (2·5 mg/d) showed no effects on the bone properties at the tibial head of ovariectomised mice. As the dosage of genistein applied in both studies was comparable after the correction of animal species, we concluded that crude isoflavone extracts such as EV extracts and Novasoy(Reference Zhang, Li and Wan14) were more superior to the pure isoflavone compound genistein in the prevention of ovariectomy-induced osteoporosis in animals. However, it should be noted that a higher dosage (1000 mg/kg) of EV is needed to achieve efficacy similar to that of Novasoy at 400 mg/kg(Reference Zhang, Li and Wan14). One possible explanation for this observation might be that the amounts of isoflavones present in the EV (10 %) and Novasoy (40 %) extracts were different. In addition, our previous in vitro data indicated that the isoflavone compounds in EV extracts, which were mainly genistein derivatives, produced more dramatic stimulation on osteoblastic proliferation, differentiation and mineralisation than genistein(Reference Zhang, Li and Yao11). Collectively, these results suggest that EV has a higher potential activity on preserving bone mass than genistein. Further studies are needed to verify the protective effects of EV extracts on bones in humans and to develop chemical methods to purify more isoflavones found in EV extracts.
To investigate the potential mechanism involved in the osteoprotective effects of EV, the mRNA expressions of an osteoblast-specific gene (cbfa1), an osteoclast-specific gene (CtsK), as well as the OPG–RANKL system in rat tibia, were studied. The present results indicated that EV extract did not alter the Cbfa 1 mRNA expression in the tibia of the ovariectomised rats, suggesting that EV extract might not stimulate bone formation in vivo. The present study showed that EV dose-dependently down-regulated mRNA expression of CtsK, suggesting that it could inhibit osteoclast-mediated bone resorption, at least partially by the inhibition of CtsK expression. This finding also explains the reduced excretion of urinary deoxypyridinoline, the degraded product of collagen, in ovariectomised rats in our previous report(Reference Zhang, Li and Lai13). OPG and RANKL play an essential role in regulating the process of osteoclastogenesis. RANKL binds to the transmembrane receptor RANK expressed on the surface of osteoclast precursors and osteoclasts, and is involved in the differentiation and fusion of osteoclast precursor cells, whereas OPG acts as a decoy receptor by blocking the interaction of RANKL with its functional receptor RANK(Reference Leibbrandt and Penninger23). The present study is the first to report that EV could up-regulate the expression of OPG and increase the OPG:RANKL ratio; thus, it inserts protective effects on bone mass by the inhibition of OPG–RANKL-regulated osteoclastogenesis.
TRAP is considered a histochemical marker of osteoclasts and is highly expressed in osteoclasts in normal condition(Reference Hayman24). Our in vitro study using RAW 264·7 cells clearly demonstrated that EV (100 μg/ml) could significantly reduce the number of TRAP-positive cells in RANKL-stimulated cells. These results indicated that EV extract could directly inhibit the process of osteoclast differentiation and maturation. They provide additional evidence to support the fact that EV extract exerts its bone-protective effects by the inhibition of the process of bone resorption. Together with our previous in vitro studies of EV on osteoblasts(Reference Zhang, Li and Yao11, Reference Li, Wang and Wong12), we conclude that EV can act on both osteoblasts and osteoclasts in vitro, but the major actions in vivo seem to be through the inhibition of bone resorption.
Taken together, the present study clearly demonstrated that EV extract is effective in improving bone properties and reducing urinary mineral excretion in oestrogen-deficient rats. Most importantly, our in vivo and in vitro results clearly demonstrated that EV extract could suppress the process of bone resorption through its direct actions on the inhibition of osteoclast-specific gene expression and the process of osteoclast formation. Further study is needed to identify the active components and compounds in EV that might account for its inhibitory activity on bone resorption. Nevertheless, the present results support the use of EV extract as an alternative regimen for the management of postmenopausal osteoporosis.
Acknowledgements
The authors would like to thank the State Key Laboratory of Chinese Medicine and Molecular Pharmacology for providing support to the present study. The study was supported by the Niche Area Research Grant from the Research Committee of The Hong Kong Polytechnic University (I-BB8N) as well as the Shenzhen-Hong Kong Innovation Circle Funding Scheme (2006).
Y. Z. and M.-S. W. developed the study protocol and the experimental design; Y. Z., Q. L., X. L. and H.-Y. W. collected the samples and conducted the experiments; Y. Z. and Q. L. analysed the data; Y. Z. wrote the draft of the manuscript with contributions from M.-S. W.; Y. Z. and M.-S. W. reviewed the final manuscript. All authors read and approved the final manuscript.
None of the authors had personal or financial conflict of interest.





