


Introduction
The snow leopard Panthera uncia is categorized as Endangered on the IUCN Red List (Jackson et al., Reference Jackson, Mallon, McCarthy, Chundaway and Habib2008). It is one of the least studied of the big cats, with little information available on its status in China in particular, which is believed to hold the greatest proportion (60%) of the global population, currently estimated to be 2,000–2,500 individuals (McCarthy & Chapron, Reference McCarthy and Chapron2003; Riordan & Shi, Reference Riordan and Shi2010). The snow leopard's range within China covers > 1 million km2 (Riordan & Shi, Reference Riordan and Shi2010), presenting a significant conservation challenge. As China's National Snow Leopard Action Plan is being developed, practical and robust techniques for assessment and monitoring of snow leopard populations are required (Snow Leopard Working Secretariat, 2013).
Developments in statistical methods have created opportunities for applying non-invasive techniques, such as camera traps, in estimating the density of snow leopards and other species and examining the influence of ecological and human covariates (Royle et al., Reference Royle, Karanth, Gopalaswamy and Kumar2009; Gopalaswamy, Reference Gopalaswamy2013; Alexander et al., Reference Alexander, Gopalaswamy, Shi and Riordan2015b). In particular, spatial mark–recapture assessments are useful for obtaining density estimates, and offer the possibility of incorporating covariates (Royle et al., Reference Royle, Karanth, Gopalaswamy and Kumar2009; Sollmann et al., Reference Sollmann, Furtado, Gardner, Hofer, Jácomo, Tôrres and Silveira2011). However, alternative methods are required when data are insufficient for mark–recapture analysis, either because the density of a species is low or photographic data are of insufficient quality for identification of individuals. We explore the use of occupancy modelling for obtaining baseline information on patterns of site use by snow leopards in cases where data are inadequate for spatial mark–recapture.
Occupancy modelling is increasingly used in monitoring programmes, especially when targeting elusive species (MacKenzie et al., Reference MacKenzie, Nichols, Lachman, Droege, Royle and Langtimm2002, Reference MacKenzie, Nichols, Royle, Pollock, Bailey and Hines2006). It represents an unbiased and cost-effective method to assess habitat use and estimate the occurrence of a species, adjusted for its detectability, supporting robust estimates of spatial distribution status and trends (MacKenzie et al., Reference MacKenzie, Nichols, Lachman, Droege, Royle and Langtimm2002; Ramesh et al., Reference Ramesh, Kalle, Sankar and Qureshi2013).
As part of a detailed study of the snow leopard population in Qilianshan National Nature Reserve, Gansu Province, we conducted a camera trap survey and applied occupancy modelling in addition to spatial mark–recapture to study the probability of site use by snow leopards. We could thus exploit even the photographs in which individuals could not be identified, which accounted for 50% of captures, mainly those taken at night or of fast-moving individuals (Alexander et al., Reference Alexander, Gopalaswamy, Shi and Riordan2015b). Our primary objective was to evaluate the influence of environmental and anthropogenic features on site-use patterns.
The selection of site-use covariates in the formulation of the candidate model set was based on local expert opinion and knowledge of snow leopard ecology. Snow leopards are usually found at elevations of 3,000–4,500 m (McCarthy & Chapron, Reference McCarthy and Chapron2003). They are agile climbers and tend to travel along mountain ridges and trails, using rocky outcrops as resting locations (Ahlborn & Jackson, Reference Ahlborn, Jackson and Freeman1988; Fox et al., Reference Fox, Sinha, Chundawat and Das1991; Jackson, Reference Jackson1996; McCarthy et al., Reference McCarthy, Fuller and Munkhtsog2005). We hypothesized that the snow leopard's preference for travelling along higher ground would be detectable even at a smaller scale, and therefore we included altitude as a covariate.
The availability of primary prey species is a key determinant of the distribution and abundance of carnivore populations (Karanth et al., Reference Karanth, Nichols, Kumar, Link and Hines2004; Sharma et al., Reference Sharma, Bhatnagar and Mishra2015). Snow leopards are opportunistic predators that exploit a wide range of prey species, with a predilection for large ungulates (Fox, Reference Fox1989; Shehzad et al., Reference Shehzad, McCarthy, Pompanon, Purevjav, Coissac, Riaz and Taberlet2012). In the Reserve, depredation of livestock by snow leopards is reported to be rare (Alexander et al., Reference Alexander, Chen, Damerell, Youkui, Hughes, Shi and Riordan2015a), and blue sheep Pseudois nayaur, which comprise the majority of large ungulate populations, are abundant. Presence of blue sheep was therefore selected as a second covariate. Finally, we hypothesized that snow leopards would tend to avoid areas where humans are active, in this case roads through the Reserve, which according to local key informants are maintained and used throughout the year (other large carnivores have been found to shun large or busy roads; Whittington et al., Reference Whittington, St. Clair and Mercer2005; Linkie et al., Reference Linkie, Chapron, Martyr, Holden and Leader-Williams2006). Distance to roads was therefore selected as a third covariate. We discuss potential methodological implications for assessments of habitat use by snow leopards across their range.
Study area
The Qilian Mountains lie on the north-eastern margin of the Tibetan Plateau. Their northern slopes, in Gansu Province, are within the transitional zone between the Tibetan Plateau (4,000–5,000 m altitude) and the Inner Mongolian Plateau (1,000–2,000 m; Wang et al., Reference Wang, Zhou, Yang and Li2003). The Qilianshan National Nature Reserve (26,530 km2), which was established in 1988 by China's State Council to protect forest and wildlife (Yang & Wang, Reference Yang and Wang2009), supports a carnivore assemblage that includes snow leopard, brown bear Ursus arctos, Eurasian lynx Lynx lynx, grey wolf Canis lupus and red fox Vulpes vulpes, and ungulates including blue sheep, Alpine musk deer Moschus alpinus and white-lipped deer Przewalskium albirostris. The study area and its surroundings are used by herders to graze livestock, principally yaks, sheep and goats. Within the study area, households kept herds of 15–530 sheep and goats and 20–140 yaks (Alexander et al., Reference Alexander, Chen, Damerell, Youkui, Hughes, Shi and Riordan2015a).
Methods
Survey design
We conducted a camera trap survey to estimate the probability of site use (ψ) and detection (p). We interpreted the parameter ψ as the intensity of use of various areas, given that the assumption of closure was invalid because of the scale of our survey (MacKenzie, Reference Mackenzie2006). Our analyses were at the scale of camera trap stations rather than home ranges, and individual snow leopards were likely to travel between stations during the survey period.
Sixty camera traps were set up across a 480 km2 study area during January–March 2013 (Fig. 1). The field area was first divided into 30 grid cells of 16 km2, and we surveyed 20 of these that were accessible (i.e. within 6 km of a road). Three camera trap stations, each consisting of a single camera unit, were set up in all but two of the grid cells surveyed. Of the remaining two cells, only two camera trap stations were set in one because of difficult terrain, and four camera trap stations were placed in a nearby cell. Camera trap stations were set up at elevations of 2,100–4,100 m, with each camera unit separated by a horizontal buffer of at least 1 km from its neighbouring cameras, except for three cameras that were separated by 310, 612 and 821 m for reasons of accessibility. The exact locations of camera trap stations were chosen based on the density of snow leopard signs and on the presence of natural pathways likely to be used by wildlife. Camera traps were mounted 30–40 cm above the ground, and the location of each camera trap station was georeferenced using a global positioning system (GPS) with a positional accuracy of < 10 m. Once the cameras were set they were left to operate 24 hours per day, with no delay between pictures, for 93 full days during the winter season. Within this period individual cameras were left unattended for 10–87 full days.
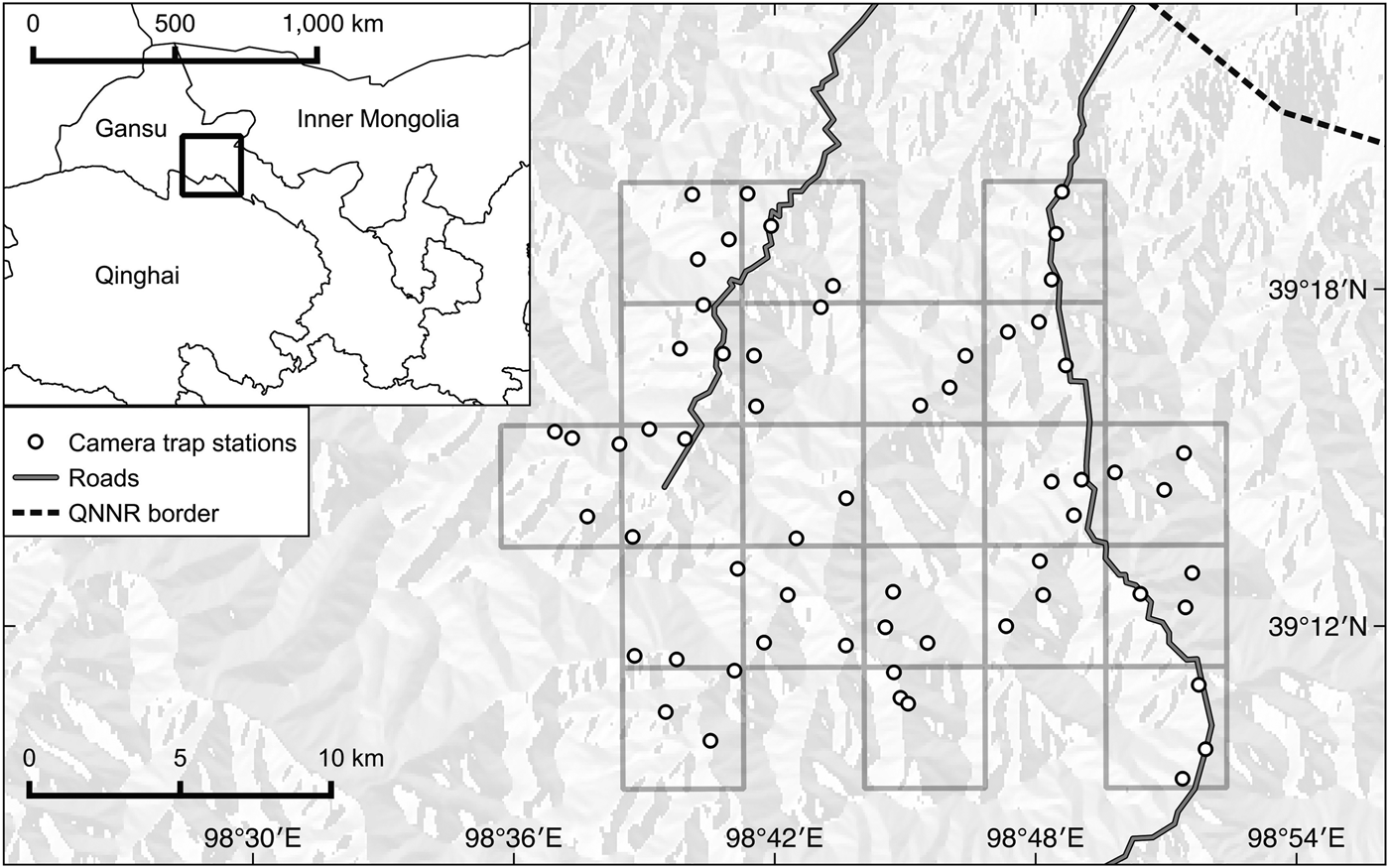
Fig. 1 Locations of camera trap stations within a grid of 16-km2 cells in Qilianshan National Nature Reserve (QNNR), Gansu Province, China.
Covariates
We collected data on potential determinants of site use by snow leopards to build our models. The first covariate, the elevation of each camera trap station, was recorded using a GPS at the time of camera trap installation. The second covariate, the presence of blue sheep, was extracted from camera trap data, with capture histories compiled as 1 for detection and 0 for non-detection at each station during a 24-hour period. Our index of presence (referred to as blue sheep presence) was the number of days on which a blue sheep was captured by a camera, divided by the total number of active trap days. Our third covariate related to human disturbance. We were unable to assess this using camera trap data of human presence because there were insufficient photographic captures. We therefore generated this covariate using remotely sensed data. During the fieldwork we observed that the study area was served by two roads, which were maintained and used throughout the winter season. Our observations were congruent with local knowledge. We digitized the roadways from Google Earth (Google Inc., Mountain View, USA) manually in QGIS v. 2.2 Valmiera (Quantum GIS Development Team, 2014) and used the NNjoin plugin to calculate the distance (in km) between each camera and the nearest road.
We gathered additional information to validate our camera trap and remotely sensed data but did not use it in the modelling process. We conducted sign surveys along 3–9 km transects (depending on accessibility) to collect data on the presence of blue sheep and human disturbance within each of the twenty 16 km2 grid cells. Signs of recent (< 1 month old) presence of blue sheep were sought along each transect. Tracks were considered to be recent if they were easily recognizable, with sharply defined edges and shape. Carcasses were considered to be recent if they were fresh or weathered, with residual signs of blood and flesh. Only signs that were unambiguously of blue sheep were recorded. We also recorded data on signs of human disturbance, as evidenced by the presence of roads, fences and intact dwellings.
Data analysis
We used PRESENCE v. 7.8 (Hines, Reference Hines2006) to run single-season occupancy models (MacKenzie et al., Reference MacKenzie, Nichols, Lachman, Droege, Royle and Langtimm2002) to estimate the probability of use, ψ. The occupancy analysis was at the scale of the camera trap station (henceforth referred to as site). Initial runs of the occupancy model in PRESENCE revealed the need to combine multiple days into a single survey occasion to increase the detection probability for each sampling period and thus reduce zero inflation in the dataset. Detections/non-detections for the first 90 days of the 93-day survey period were combined over 5-day periods for each camera trap station, resulting in 18 sampling occasions. Data from the last 3 days were discarded. The 5-day period provided the best balance between the number of sampling occasions and the probability of detection in any sampling occasion. Detection was defined as the capture of an individual snow leopard by a single camera trap. Snow leopard detection histories were coded as 1 for detection and 0 for non-detection during each 5-day period. Photographs of more than one individual were counted as a single detection.
To reduce the number of candidate occupancy models we adopted a two-stage model-building approach. To prepare for the analysis all covariates were standardized to z scores. Using the Pearson correlation test we explored relationships amongst covariates, to avoid problems with multicollinearity. We considered variables to be highly correlated when correlation coefficients were > 0.7 in magnitude (Dormann et al., Reference Dormann, Elith, Bacher, Buchmann, Carl and Carré2013). In the first stage of analysis we defined a global model for ψ (elevation + blue sheep presence + distance to roads; Burnham & Anderson, Reference Burnham and Anderson2002) and then held the global ψ constant as we developed the most suitable model for detection probability, p. In the second stage we used the best detection model (based on Akaike's information criterion, AIC) from the first stage and ran our candidate set of models to test the effects of covariates on ψ (Royle & Nichols, Reference Royle and Nichols2003). To identify the best ψ model we fitted all combinations of the three covariates in our hypotheses relating to site use (elevation, presence of blue sheep, and distance to roads).
We assessed model fit by running bootstrap goodness-of-fit tests for the global standard occupancy model (n = 10,000 bootstrap samples), using the overdispersion parameter c-hat calculated in PRESENCE. Values of c-hat > 1 indicate there is more variation in the observed data than expected and the associated AIC values should be adjusted to compare models (MacKenzie & Bailey, Reference MacKenzie and Bailey2004). Values of c-hat < 1 indicate less variation than expected and do not pose a problem (Burnham & Anderson, Reference Burnham and Anderson2002), and can be addressed in PRESENCE by setting c-hat to 1.
The information-theoretic approach was used to explore the most appropriate models from the suite of occupancy and detection models, drawing on the least number of parameters considered necessary (Johnson & Omland, Reference Johnson and Omland2004). AIC performs poorly if there are too many parameters in relation to the effective sample size (Burnham & Anderson, Reference Burnham and Anderson2002). We used AICc (AIC adjusted for small sample size) to rank models because of our small number of camera traps (ratio of sample size (n) to parameters (k) < 40).
We assessed the relative importance of each covariate by summing AICc weights of all models that included the covariate. The beta coefficients of covariates (β) reveal the effect size and direction of influence. We used model averaging to calculate weighted average estimates of ψ and p for each model, following Burnham & Anderson (Reference Burnham and Anderson2002). Standard error was calculated using parametric bootstrapping, following Karanth et al. (Reference Karanth, Gopalaswamy, Kumar, Vaidyanathan, Nichols and MacKenzie2011). The model-averaged coefficients (ψ and p) for each individual camera trap site and across all sites, and their unconditional standard errors, were computed following Burnham & Anderson (Reference Burnham and Anderson2002).
Finally, we used a simulation study to investigate the bias and precision of our ψ estimate (Bailey et al., Reference Bailey, Hines, Nichols and MacKenzie2007). Using our final parameter estimates for ψ and p, 1,000 simulation replicates were generated in GENPRES (Bailey et al., Reference Bailey, Hines, Nichols and MacKenzie2007) with the single-season model (MacKenzie et al., Reference MacKenzie, Nichols, Lachman, Droege, Royle and Langtimm2002). We simulated camera-trap sample sizes of 60, 100 and 150. We repeated the simulation using detection probabilities of 0.20 and 0.30, with a sample size of 60 camera traps. For each simulation we approximated bias and precision by calculating the root mean square error. A lower root mean square error indicates a higher quality parameter estimate.
Results
We recorded 81 captures of snow leopards. Three of these were discarded because the same individual was captured by the same camera trap within an interval of a few hours. To maintain a standard duration for each trap-day two more captures were discarded because they were made before 24 hours of camera-trap pickup. We therefore recorded 76 captures over 2,906 trap-days, representing a mean capture success of 2.62 captures per 100 trap-days. Snow leopards were recorded by 30 camera traps (50% naive occupancy). Sympatric mammal species (Eurasian lynx, red fox and grey wolf) were also recorded, although less frequently, as were herds of yaks, generally accompanied by herders on horseback.
Covariates were not strongly co-dependent (all Pearson correlation coefficients < 0.41). The camera traps were set up at various altitudes (mean = 3,166 ± SD 465 m, range 2,082–4,092 m) and various distances from roads (mean = 2.97 ± SD 2.65 km, range 0–8.50 km). Blue sheep were detected on 46 of 2,906 trap-days, at a total of 19 (32%) camera trap stations, and tended to occur in groups. Our sign surveys covered a total of 100 km across all 20 grid cells (mean = 5 ± SD 1.49 km per grid cell, range 3–9 km). Signs of blue sheep were found in all grid cells. Signs of human activity (fences, roads and any intact dwelling) were observed in 55% of grid cells.
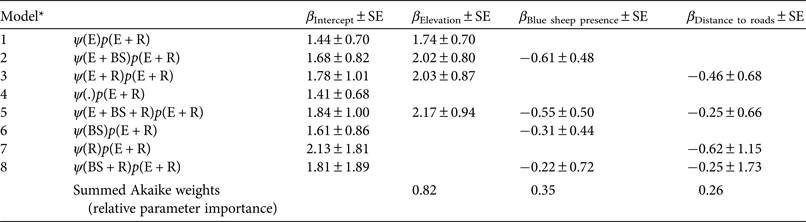
The goodness of fit test for the global standard occupancy model indicated no overdispersion, with c = 0.67, p = 0.28. In the first stage of modelling, snow leopard detection (p) was best explained by the model that included elevation and distance to roads (AICc weight = 0.38). The probability of detecting snow leopards decreased with elevation and closer to roads (β = –0.47 ± SE 0.25 and 0.58 ± SE 0.16, respectively), and therefore all further analyses included distance to roads and elevation in modelling detection. In the second stage we tested eight models to assess ψ (Table 1). Elevation had the strongest influence on probability of site use by snow leopards (summed AICc weight = 0.82), occurring in all three top models. Based on the β estimates, snow leopards were more likely to use areas at higher elevations (Table 2). Presence of blue sheep and distance to roads had a lesser influence on the probability of site use by snow leopards (summed AICc weight = 0.35 and 0.26, respectively). Both covariates had a small negative effect on probability of site use, and the predictive power of these two covariates should be interpreted with caution given their large standard errors (Table 2). The model-averaged estimate of ψ at individual camera trap locations was 0.21–0.97. The final estimates of probability of site use (ψ) and detection (p) averaged over all sites were 0.75 ± SE 0.10 and 0.15 ± SE 0.03, respectively (Table 3), indicating that the naïve estimate of site use underestimated occupancy by 25% (Fig. 2).

Fig. 2 Probability of site use by snow leopards Panthera uncia, as measured by camera traps in Qilianshan National Nature Reserve (Fig. 1); each cell is 16 km2. (a) Naive estimates from a presence vs absence approach. (b) Mean estimated probabilities of site use.
Table 1 Summary of model selection results indicating the role of covariates in determining probabilities of snow leopard Panthera uncia detection and site use (n = 60 sites), with Akaike's information criterion adjusted for small sample size (AICc), change in AICc (ΔAICc), Akaike weight, model likelihood, no. of parameters (k), and 2log-likelihood (LL).

* Covariates: elevation, E; presence of blue sheep Pseudois nayaur, BS; distance to roads, R
Table 2 Estimates of β coefficient values and summed Akaike weights for covariates that were hypothesized to influence site use by snow leopards in Qilianshan National Nature Reserve (Fig. 1).
* Covariates: elevation, E; presence of blue sheep, BS; distance to roads, R
Table 3 Summary of model-averaged parameter estimates of probability of site use (ψ) and detection (
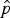 $\hat p$
) for snow leopards in Qilianshan National Nature Reserve (Fig. 1).
$\hat p$
) for snow leopards in Qilianshan National Nature Reserve (Fig. 1).

1 Covariates: elevation, E; presence of blue sheep, BS; distance to roads, R
2 Model-averaged estimates across all models with unconditional standard errors
Our simulation based on 60 camera trap sites estimated a probability of site use of 0.89 ± SE 0.15 and indicated that our model estimates of occupancy were not optimal (root mean square error for site use 0.20). In the simulations with increased numbers of camera trap sites (100 and 150), lower root mean square errors were achieved (0.09 and 0.06, respectively). The quality of estimates was improved substantially when we simulated higher detection probabilities of 0.20 and 0.30 with 60 camera trap sites (root mean square errors of 0.13 and 0.06, respectively).
Discussion
Determinants of site use by snow leopards
Threats to snow leopards are often context-specific and should be appraised at the local level. We set out to explore methods for assessing the determinants of snow leopard presence at a small scale, and to exploit an alternative method of assessing such determinants in situations when not all of the sightings are suitable for mark–recapture methods.
We observed that site use by snow leopards was not strongly associated with the presence of blue sheep or roads, contrary to our initial hypotheses. Prey is known to be a key determinant of the presence of snow leopards (Sharma et al., Reference Sharma, Bhatnagar and Mishra2015) and other carnivores (Barber-Meyer et al., Reference Barber-Meyer, Jnawali, Karki, Khanal, Lohani and Long2013). A study in Himachal Pradesh, India, confirmed that the abundance of wild prey (blue sheep and ibex Capra sibirica) was the main explanatory factor for site use by snow leopards within an area of 4,000 km2 (Sharma et al., Reference Sharma, Bhatnagar and Mishra2015). In our smaller study, the presence of blue sheep was detected widely across the study area, with no distinct geographical concentration. The sign surveys confirmed the presence of blue sheep in all 20 grid cells. This observed lack of heterogeneity may have contributed to the weak association detected, coupled with the high probability of site use by snow leopards across the landscape surveyed. Blue sheep were clearly present within the territory frequented by snow leopards.
There are a number of possible reasons why we found little empirical support for the presence of roads as a determinant of site use by snow leopards. In the study area, roads represented foci of human activity. In the winter months, however, vehicle access to the area is constrained by adverse weather and overflow of riverine ice. One of the roads led to a dam construction site (covering an area of c. 1.6 km2), and construction work was interrupted during the winter months. Furthermore, human activity was not limited to roads, and the camera traps provided evidence of people herding yaks across the landscape. Other traces of human activity included simple habitations, which may also be associated with livestock rearing and the accompanying seasonal human presence. All these types of human activity are likely to be greater outside the winter season. We believe that the presence of roads remains a useful indicator of human activity in this remote area and is worth exploring as a potential determinant of site use by snow leopards in different seasons.
Most importantly, however, the scale of our analysis may well be masking the true effects of the covariates indicating prey presence and human disturbance (Sunarto et al., Reference Sunarto, Kelly, Parakkasi, Klenzendorf, Septayuda and Kurniawan2012; Srivathsa et al., Reference Srivathsa, Karanth, Jathanna, Kumar and Karanth2014). Our survey results suggest that snow leopards use a large proportion (75%) of the area. Again, this lack of heterogeneity makes it difficult to establish clear relationships between site use and the selected covariates. The lack of spatial independence among adjacent sampling units may further weaken inferences, given that we did not account for spatial autocorrelation in our analysis. Three of our camera traps were < 1 km apart. More work is needed to explore the determinants of site use by snow leopards at various geographical scales.
In contrast, altitude was found to influence site use by snow leopards at this scale. Snow leopards clearly favoured areas at higher elevations, and camera traps often documented individuals travelling along ridgelines. Qilianshan National Nature Reserve is on the boundary between Gansu's inhabited Gobi desert and Qinghai's mountainous Tibetan Plateau. Towards the south-western parts of our study site the elevation increases sharply, from 2,436 to 4,387 m. Elevation may therefore be correlated with increasing remoteness and diminishing human interference. However, our findings refer to the areas that were accessible to the survey team and should not be generalized to more remote areas without further verification.
Methodological considerations
Snow leopards are notoriously elusive, and the failure to detect them when they are present is a common source of survey bias (Linkie et al., Reference Linkie, Dinata, Nugroho and Haidir2007). In addition, detectability may vary with site or survey characteristics (MacKenzie et al., Reference MacKenzie, Nichols, Lachman, Droege, Royle and Langtimm2002, Reference MacKenzie, Nichols, Royle, Pollock, Bailey and Hines2006). Few previous assessments of snow leopard distribution and site use have modelled or estimated detection probabilities explicitly, which may have led to underestimates of spatial distributions.
Our approach sought to account for these two sources of bias (imperfect detection and spatial variation in site characteristics), which can otherwise lead to unreliable estimates. Our simulation results may inform the design of studies where detection of snow leopards is particularly low (p = 0.15), to increase precision and minimize bias. Increasing the number of camera traps from 60 to 100–150 at the same density would improve the performance of the occupancy models, lowering the root mean square error substantially. However, such an investment in camera traps and field effort may be difficult for local teams with limited resources to achieve. Alternatively, measures could be taken to increase detection probability. The simulation results indicate that estimates of site use would be improved by increasing detection probability to 0.30, while keeping the number of camera traps constant (n = 60). This could be achieved using sign surveys instead of camera traps, as has been shown in research on other carnivores (Steinmetz et al., Reference Steinmetz, Seuaturien and Chutipong2013). The application of multiple detection methods (Nichols et al., Reference Nichols, Bailey, O‘Connell, Talancy, Campbell Grant and Gilbert2008) may also be worth considering. Sign surveys have the logistical advantage of covering large areas, with lower field efforts compared to camera trap supervision and maintenance. However, care is needed to minimize the misattribution of scats (Janečka et al., Reference Janečka, Jackson, Yuquang, Diqiang, Munkhtsog, Buckley-Beason and Murphy2008) and other signs (Miller et al., Reference Miller, Nichols, McClintock, Campbell Grant, Bailey and Weir2011) to snow leopards.
Conclusion
The appraisal of threats to snow leopards can provide important information for strengthening local conservation. Expert opinion is often the main source of such information (Snow Leopard Network, 2014). Verification or updates using field data together with robust analytical methods are required to capture the diversity of local settings. One of the purposes of our study was to test simple and robust approaches for examining determinants of site use by snow leopards, which could potentially be used by local field staff in protected areas. Caution is required when carrying out assessments at the scale presented here, as limited inferences yield weak information for policy decision making. This points to the investment case for larger scale efforts to assess threats, to gain a clearer understanding of the determinants of snow leopard distribution and site use in a given area, and how they may vary across the region.
China, with its network of nature reserves, has the advantage of being able to mobilize a significant resource of human capital, represented by local teams, in efforts to conserve the snow leopard. For this resource to be harnessed, however, there is a need for methodological approaches that are replicable, user friendly and robust. Our findings present clear advancement but further efforts are needed in the design and appraisal of such methods.
Acknowledgements
We acknowledge the support of China's State Forestry Department, and the National Natural Science Foundation of China (Grant No. 31470567). JA acknowledges support from the National Geographic Young Explorers Grant. PR thanks the Starr Foundation for support during the undertaking of this work. We thank Qilianshan National Nature Reserve, and Wang Youkui and A. Chen for their help and support in the field in Gansu Province. JA also thanks Wang Minghai, Wu Qian and Joelene Hughes for their support during fieldwork.
Biographical sketches
Justine Shanti Alexander is studying the conservation of snow leopards in high-elevation areas of China. Shi Kun focuses on translating academic research into conservation through training local wildlife managers, building a collaborative network and applied research. Lucy Tallents creates innovative online and face-to-face training in research and monitoring techniques to build the capacity of in-country conservation practitioners. Philip Riordan has worked in the UK, China and Africa, focusing on top predators, their ecosystems and their interactions with people.





